
本シリーズは、昨年(2009年)の、
1月29日アップの
色の話 その1

2月16日アップの
色の話 その2

3月18日アップの
色の話 その3−付 日本産ハンミョウ科チェックリスト−
の続編です。
当時は、引き続き、その4もアップの予定でしたが、Knisley & Schults, 1997論文の入手と理解に手間取り、1年遅れになったものです。
このページからご覧の方は、併せて、色の話 その1から順に覗いて頂く方が、理解しやすいと思います。
さて、4回目にして、やっと、前回お約束の北アメリカ産を紹介します。
40年近く前に、北アメリカのテキサス州のほぼ中央部、名前もずばり、ミッドランドという町に、一夏、ホームステイをしたことがあります。
この町の住宅地周辺の砂漠(丈の低い草地が続く半砂漠・ステップと言うべきか)で採集したハンミョウを紹介します。
なお、北アメリカ産については、全て、堀 道雄京都大学教授(以下、堀さんと省略します)に、写真同定により、名前を教えていただきました。
{上:Midland郊外から中心部を望む、下:赤茶けた地表に静止するCylindera lemniscata (Leconte)}


この地は、メキシコ国境までほど近く、緯度も九州南部程度の場所で、亜熱帯の生き物も見られ、庭の花にはハチドリも飛来しました。
最初の種{Cylindera lemniscata (Leconte)}は、赤銅色に輝く綺麗な種で、そんな、半砂漠の赤茶けた砂地の地表を走り回っていました。
(左:背面, 右:上翅)


本種では、凹部が波形に横に繋がり、点刻の底には盃刻は見られません。
また、凸部も畝状に横に繋がり、こちらは印刻で密に被われ、そのうちの一部は、印刻の編み目の中央が凹み、中には、かなり深さを増して盃刻に近づいた構造も見られます。
「盃刻の最初は、こんなだったのかな?」と、思える感じです。
凸部は美しい赤銅色ですが、凹部の底は金色に光っています。
次いで、Ellipsoptera cuprascens (Leconte)です。
こちらは、Midlandから100kmくらい走った大きな渓谷で、谷間の裸地を走り回っていた種です。本種もカンカン照りの河原で、元気に活動していました。
(上:背面, 下:上翅)


体長と斑紋パターンはエリザハンミョウに良く似ていますが、白紋はより太くて広く、カワラハンミョウのように、金属光沢部分はかなり縮小しています。金属光沢のある凸部の色はやや暗い赤銅色で、点刻の底は緑がかった金色です。
やや不明瞭ですが、白紋も凸部も、網目状になった印刻に混じって、一部にごくごく浅い盃刻様の構造が見えます。点刻の底部には、印刻も盃刻様の構造も見られないのは、前種と同じです。
この2種の状態は、今まさに、印刻(凸部の)が盃刻に変化しつつある状態のように思えます。
もちろん、このミッドランドでも、コハンミョウにも似た、ちゃんとした盃刻を持つ普通タイプの地表性ハンミョウも何種か見られましたが、そちらの盃刻の構造は日本産と変わらないので割愛します。
次に、特筆すべき種として、Megacephala carolina (Linne)を紹介します。
この種は18mm程度とやや大型で、体形や色からは、とても普通のハンミョウの様な感じがしません。
(上:背面, 下:上翅)

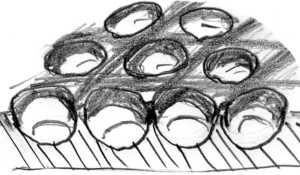
この個体は、牧場にある牛の水飲み場近くの、やや湿った石の下に潜んでいました。石の下から、こんな極彩色の、それも変なハンミョウが出てきて、どぎまぎしたのを覚えています。
上翅表面には、細かく弱い印刻があるだけで、かなり光沢が強く、盃刻はありません。日中の暑いときは、石下などに潜んで休憩しているようなので、盃刻までは必要無いのかも知れませんね。
堀さんに確かめたところ、この属は、南半球に広く分布する属で、中でも本種が含まれる亜属Tetracaは、南米で大繁栄(約90種)しているグループだそうです。
北アメリカでもテキサスまで来ると、かなり、南米の匂いがしてくるようで、滞在中、ツルギタテハやハチドリなどに、そのような片鱗を感じました。
この属は夜行性とまではいかず、南米では主に林床性であり、一方、オープンな所、特に砂漠などでは夜行性および薄暮性(夕方に活動する)のようです。曇りの日は日中でも活動するとされています。この属には後翅が短く飛べない種も多いのですが、本種は良く飛び、灯火にも飛来するそうです。
北米には、この他にも本属に含まれる種がもう一種いるそうです。
さて、北アメリカ産の最後に、Amblycheila cylindriormis (Say)という、さらに変わったハンミョウを紹介します。
(上:背面, 中:頭部, 下:上翅)



体長は30mmほどと巨大で、足は短く、目も小さく、前胸と頭は黒、上翅は黒褐色です。
上翅には当然、印刻があるだけで、金属光沢も、盃刻も、ありません。
この種は姿から想像できるように、夜行性のようです。
昼間はヒョウタンゴミムシ同様、砂の奥に潜っていて、夜に活動し、徘徊しながら捕食するのでしょうか?
(堀さんの解説によると、「日中は哺乳類の掘った穴または石の下にいて、夜活動する。後翅は短く、飛べない。鞘翅も開けない。」ということだそうです。)
堀さんにご教示いただくまで、この種を、私は多分オサムシ科からはやや類縁が離れた、○○ゴミムシ科などと言う、アメリカ固有の科のゴミムシと思っていました。
北米の砂漠にも多いヒョウタンゴミムシともちょっと違い、オサムシとも違い、変な奴とは思っていましたが、まさか、これが、ハンミョウの中に入れられているとは思いもよりませんでした。同属種が合衆国西南部とメキシコに計7種分布するそうです。
残念ながら、砂に埋もれた死骸をいくつか拾っただけで、生きて活動している姿は見ていないので、どんな生活をしているのか解りません。
かの地は、気候的には砂漠で、滞在した6週間のうち、にわか雨を2回ほど見ただけです。
地表もカラカラに乾いていて、地表性のゴミムシやゴミムシダマシ、庭木に付いたカミキリなどの死骸が、まったく腐らず乾燥標本状態で砂に埋もれていて、それらを結構拾うことが出来ました。
ゴミムシからハンミョウに分化した最初が、こんな感じと思われますが、この種がハンミョウの範疇に入るとしても、最初の3種などの直接の祖先ではないでしょう。
だいぶ、系統的には違ったもののように思われます。
(しかし、堀さんの解説によると、「Amblycheila はMegacephala族なので、Tetrachとは近く、むしろMegacephalaとCicindela族の間にはかなりのギャップがある。」ということだそうです。
AmblycheilaとMegacephalaには盃刻が無く、Cicindelaには盃刻が有る、という点においては、そのことは支持できますが・・・。)
このようなものから、どんな風にして、昼行性のハンミョウが進化してきたのでしょうか?
なお、以下の論考に堀さんからも助言をいただきましたが、今坂の考えの細部まで支持されているわけではないことを、お断りしておきます。
<盃刻の構造>
前回まで3回に渡って見てきたように、日本産の地表性のハンミョウには、殆どの場合、上翅表面に盃刻が見られました。
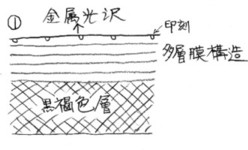
盃刻を含む上翅断面を、私の考える模式図として描いたのが次の図です。
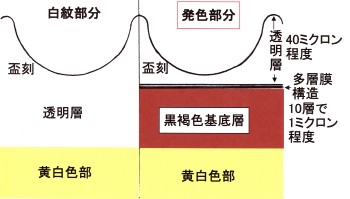
鞘翅は、裏面から見ると、黄白色に見えます。
この部分は、羽の強度と柔軟性を増すために、黄白色の繊維質のもので裏打ち(コーティング)をされています。
白紋の部分は、その黄白色部が、表から直接透けて見えているものと理解されます。
ですから、白紋の部分は、背面側には透明な層が、基底の黄白色層の上に乗っていると考えられます。
白紋表面には盃刻の構造がありますが金属光沢は見られず、このことから(表面の盃様のへこみ部分としての)盃刻の構造自体に発色する機能は無い、と考えています。
次に、コニワハンミョウの♀などに見られる鏡紋の部分ですが、ここは黒褐色で表面はツヤがあります。
鏡紋の表面を傾けて光を当てると、角度により盃刻部分とは異なる油膜状の光が見えますので、鏡紋部分では、黄白色層の上に黒褐色層が乗り、さらに、表面には透明な薄い層が有るのだろうと思います。
さらに、前2者以外の、普通に盃刻が見られる部分は、凸部も凹部も含めて、下層から黄白色層の上に、黒褐色層が重ねられ、その上に、金属光沢を発する層、そして、表面に盃刻という重なりになっているのだろうと思います。
<甲虫の金属光沢>
さて、甲虫の一般的な金属光沢については、従来、どのような構造から生み出されているのか、少し前まで、色んな意見はあっても、定説がありませんでした。
しかし、欧米では、20年くらい前には、かなり解明が進んでいたようです。
日本語の解説として手頃に読めるものとしては、琵琶湖博物館から発行された「オサムシ」に寄稿されている吉岡さんの文章があります。
吉岡伸也 (2008) オサムシの輝く翅の秘密. 琵琶湖博物館ポピュラーサイエンスシリーズ オサムシ,: 42-48. 八坂書房.
この本から、以下のように、図を引用させて頂きます。
(上:オサムシ表紙、下:文章タイトル)


(画面左側の縞→多層膜構造)
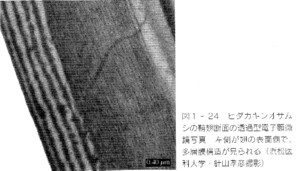
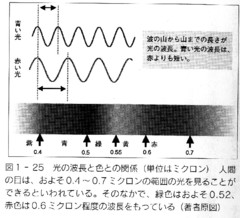
吉岡さんの解説によりますと、甲虫の金属光沢は、上翅表面にある薄い透明な膜の多重構造によって発現されているとのことです。
0.1ミクロン程度の薄い膜の表面と裏面で、それぞれ、光の反射が起こり、薄膜の厚さによって、表面からの反射光と、裏面からの反射光が、強調したり打ち消し合ったりして、特定の波長の色だけが選択的に反射されることにより、人の目からは一定の色の金属光沢と認識されるそうです。
より薄い膜では波長の短い青い光が、より厚い膜では波長の長い赤い光が反射されるようです。
甲虫では、この薄膜が8〜10層程度重なって、それぞれの膜で光を反射し、それが複合されて、かなり、複雑な金属光沢が形成されているようです。
詳細は、上記文献の解説を参照下さい。
前項での、金属光沢を発する層、というのが、この多層膜構造に当たると考えています。
<盃刻の形成>
北アメリカ産のCylindera lemniscataとEllipsoptera cuprascensは、砂漠地方のカンカン照りの砂地で生活していますが、上翅表面にはそれぞれ、強い金属光沢や、かなり広い白紋を持っています。
この、鮮やかな金属光沢や白紋で、太陽光のかなり多くの光を(それも熱を多く含む赤色系の光を)反射しているので、わざわざ盃刻構造までは必要ないのかも知れません。
しかし、実際には、一部は印刻の真ん中をちょこっと凹まして、表面積をちょっと増やし、盃刻様に変化しかけていますので、さらに、熱を逃がす効果を強化したいと思っているのかも知れません。
さらに暑い地域で生活するなら、もっと放熱効果を高める必要があります。
その為に、それらの個体群は、さらに印刻の真ん中を凹まして、盃刻を完成させたのでしょう。
当然、引き続き、普通の金属光沢は持ったままだったはずです。
さて、Cylindera lemniscataの、あの赤銅色の色彩では、赤茶けた地表で生活する場合はそれほど目立たないと思いますが(というより、日中の眩しいほどの陽光の中では逆に乱反射もありよく見えませんでした)、黒っぽい地面の上では、ひどく目立つことになると思います。
そんな場合、なんとか、体色を暗い色彩に見せることが有利に働くに違い有りません。
しかも、放熱効果のある金属光沢を保ったままなら、なおさら有効です。
ちょっとした工夫で(この、ちょっとした、というのが重要です。大幅な変更は簡単にはできませんので)、彼等はそれを実現しています。
上翅の金属光沢を、違った色の細かいパーツに分割して、補色関係になるように、隣り合わせに配置すれば良いわけです。
このことを実現したのが、日本産を含む地表性ハンミョウの一群、広義のCicindelaだと考えたわけです。
つまり、私は、北アメリカの夜行性のAmblycheila cylindriormis、薄暮性のMegacephala carolina、さらに昼行性の2種や日本産の表面構造から推測して、ハンミョウ上翅の構造の変化を次のように推定したわけです。
まず、
1. ハンミョウの最も原始的な夜行性の初期の段階では、体色表面は黒〜褐色で、金属光沢は無かった。
2. さらに、やや発展した薄暮性のMegacephalaくらいになると金属光沢を獲得した。
3. その後、表皮に普遍的に見られる印刻の、各編み目の中央をへこますことによって、盃刻を獲得した。
と3段階に考えているわけです。
可能性としては、逆に、盃刻を獲得してから、その下に金属光沢(多層膜構造)を作ることも否定は出来ませんが、その順序での獲得は、必然性も含めて、構造的にはかなり難しいだろうと思います。
さらに、次の段階の、反対色の配置による補色配列の獲得ということですが、これは比較的簡単に実現できたのではないかと思います。
このことには、先ほどの吉岡さんの解説がヒントになります。
簡単に言うと、多層膜構造では、色は単に、薄膜の厚さだけで決まります。それなら、この厚さを変えれば、色を変えることが出来るわけです。
先に示したように、日本産の盃刻を持つ大部分のハンミョウでは、上翅凸部〜平坦部は赤銅色に光っていました。
一方、凹部の底面では青紫色、そして、両者の中間では黄金色〜緑色であるのも、共通です。
先に紹介したハンミョウ上翅の断面図を念頭に置いて、その前の状態を想像してみて下さい。
まず、金属光沢がある上翅(表層部には多層膜構造)を考えます。表面には網目状の印刻を備えています。
なお、多層膜構造の部分は、本来は、盃刻のサイズに比べて、1/50〜1/100程度の厚さですが、図として見やすいように、100倍以上に拡大し厚みを拡げて書いています。
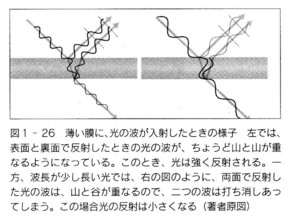
次いで、印刻の中央部を凹まして、この多層膜の表面の一部に、熱放出のために盃刻構造を作ります。

この時点では、まだ、Cylindera lemniscataのように、鞘翅のほぼ全体が、同一の光を放っていた可能性が強いと思います。
さらに、盃刻の下の上翅表層に、盃刻よりずっと大きなゆるやかな凹凸を作ります。
この場合、断面図を想像すると、凹んだ部分では、多層膜構造の下縁は、その基底の黒褐色層に食い込んで凹んだ形になると思います。

次に、この、黒褐色層を元の均一な厚さに戻してやると、凹んだ部分では多層膜構造全体の厚みを減じるしかありません。
そうすると、必然的に、この部分では一層一層の薄膜の厚みは薄くなります。
結局、凹部では薄くなった分、そこで反射する波長も短くなって青紫色にならざるをえません。
また、当然、中間にある斜面部は、中間の厚さで黄金色〜緑色になり、平坦部では元のままの赤銅色です。
これで、Cicindela達の色のモザイク配列は完成するわけです。

盃刻と金属光沢を持つ種が、全体的になだらかな凹凸をつけ、基底の黒褐色層を元の均一な厚さに戻すと、自然に、モザイク様の点描画が作れるわけです。
後は、この凹凸の配置と密度で、どんな色にも対応できます。必要な時には、生活場所に合わせて色を変え、隠蔽色に利用したのだと考えています。
繰り返しますが、
1. まず、光と熱を反射する金属光沢があって、
2. 次に、効率よく熱を逃がす盃刻構造の獲得、
3. さらに表面にゆるやかな凹凸を作ることによる色のモザイク配置、
4. それを利用した地表色に似せた隠蔽効果の獲得、
この順で進化していったのではないかと考えています。
それから、言い忘れていましたが、金属光沢の発色には基底の黒褐色部の存在が欠かせません。
この部分が、白紋の様に白色、あるいは透明のままだと、それぞれの薄膜であちこちに散乱した色んな波長の光を全て反射して、特定の波長の光が目立つことを弱めてしまいます。
黒褐色部で、余分な光を吸収することによって、特定の光だけを際だたせて反射させることが出来るわけです。
最初、ハンミョウ表面のあまりにも独創的な形の構造物である盃刻に目が行き、モザイク様の金属光沢が、この構造に起因すると考えました。
しかし、発色の機能や構造を細かく見ていくと、根本的な色のモザイクは、むしろ、本来の金属光沢を担っている多層膜構造と、表面のなだらかな凹凸、そして、黒褐色の基底層の、三者の共同関係で成り立っていることが理解されました。
発色そのものには、表層の盃刻構造はかかわっていない、というのが、現在の私の考えです。
盃刻の枠組みは、反射された金属光沢を細かく細分化し、あるいは、盃の縁の部分の立体的な反射によって色の輪郭を作り、ちょうど、デジタルテレビで、色点の周りに黒の縁取りをして、画面をくっきりさせるというような、そんな効果を担っているのではないかと思います。
以下の文章中では、表層のくぼみを盃刻あるいは盃刻構造と呼び、ハンミョウ上翅の発色構造の全体である盃刻+多層膜構造+黒褐色基底部の全てを含めて呼ぶ際は、盃刻発色構造という言葉で呼ぶことにします。
<Tom Schultz (1991) による盃刻発色構造の解説>
この報告を書き始めた頃からその後しばらくの間、私自身の意識としては、盃刻発色構造は自ら発見したつもりでいました。
しかし、実は、既にアメリカのTom Schultz (1991)が、ハンミョウ上翅にあるこの構造について紹介しているということを、堀さんに教えていただきました。
Schultz, T. (1991) Tiger Hunt. Natural History, 5: 38-44.
さっそく、お願いして論文を送ってもらい、読んでみると、当然、私がその1で示した模式図より詳細な構造が解説されていました。
以下にその図を引用します。
きれいな、ビーズ模様の写真も同じように掲載され、私同様、白紋の部分にある盃刻も見えるように配置されています。
ハンミョウの盃刻発色構造による色のモザイク模様は、私が知らなかっただけで、20年程前には既に報告され、研究者間では周知の事実であったようです。

彼は、盃刻を単にpit(穴)と呼んで、その形状を「ワッフルに似ている」と表現しています。
彼が甘〜いお菓子を連想し、それほどお酒を飲まない私が、お酒の盃を連想したのは、我ながら、ちょっと、可笑しい気がします。
彼は甘党なのでしょうか?
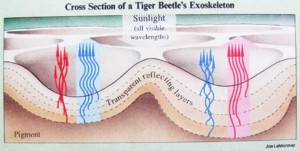
彼は、盃刻の浅層構造それ自体の中に、金属光沢を反射させる多層膜構造が組み込まれているように、模式図を描いています。
しかし、既に述べたように、白紋上の盃刻からは金属光沢が観察されないことを理由に、私は、盃刻自体は単純な透明層で、多層膜構造は盃刻の下に、別の層として存在すると考えています。
彼は、盃刻発色構造の色のモザイク配列を、新印象派のスーラの点描画を引き合いに出して説明し、いくつかの単純な色の配列具合で、どんな地面の色にも、体色を似せることが出来ることを強調しています。
生態の専門家である彼にとっては、体色をいかにして変化させて、周囲の色に紛れ込ませるか、ということの解明が最大の関心事だったのでしょう。

さらに、堀さんのご教示によると、Schultz等はその後に書いた以下の論文の中で、さらに発展させた議論をしているそうです。
Knisley & Schultz (1997) The biology of tiger beetles and a guide to the species of south Atlantic states.
私自身は、論文を読んでおりませんので、論文に目を通された堀さんの感想を、以下に引用します。
「もともと、彼(Schultz)は、ハンミョウの体色の生態的意味を追究している研究者ですが、この本ではその集大成ともいえるまとめをしており、体色の意味を、温度適応と敵からの保護(目立たないようにする効果)とのバランスで現れる適応と、位置づけています。
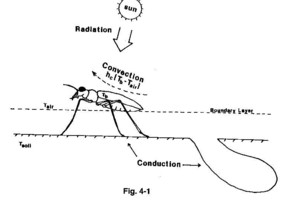
図4-1、4-2では、体温調節に関わる要素が示されていますが、radiation(太陽放射)をいかに吸収し、あるいは反射するかに注目しています。
そして周囲の温度から少し高めの体温を積極的に維持していることが示されています。
その測定のために、微少な電極の温度計を使っているようです。
Schultzの扱っている種は、どちらかといえば冷温帯の種類が主で、熱を逃がすという観点が弱いかもしれません。」
(Knisley & Schultz, 1997 による図4-1より引用)
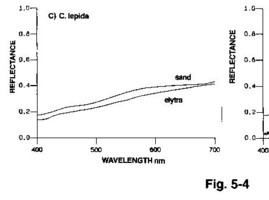
「別の図(Fig.5-4)では、エリトラからの反射光のスペクトル(分光放射計という器械で測定)がいかに周囲の反射スペクトルとマッチしているか、つまり虫体が目立たなくなっているかが示されており、説得力があります。」
(Knisley & Schultz, 1997 による図5-4より引用)
「これはエリトラ全体からの反射についてであり、盃刻の機能そのものを調べているわけではありませんが、印象派の点描などの言及からもわかるように、盃刻の集合効果を念頭には置いているでしょう。
気温適応については、盃刻が関与しているかどうかについての言及は見つけることができませんでした。
私も盃刻という微小な構造がどこまで温度を下げる効果を持つかはよく解りません。
光の反射だけでなく体表面の空気の動きも関わってくるでしょう。
ですが、熱の吸収なり反射なりが、いきなり体色に出ないようになっているという機能的意味については今坂さんの意見に賛成です。
これについてはSchultzも同意見ではないでしょうか。
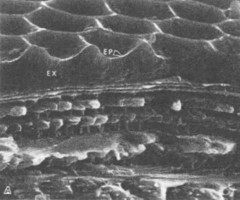
盃刻とエリトラの断面構造については、走査電顕で調べており(Fig. 1-1)、これは今坂さんの考えと合致している構造です。
ただ、色の発色に基底層が関わっているかどうかは読み取れませんでした。」
堀さんの解説と資料から、今坂が感じたのは以下の点です。
この論文でSchultz等は、盃刻発色構造の機能として、光と熱を吸収する効果を考えているようです。
これは、堀さんも指摘されているように、「Schultzの扱っている種はどちらかといえば冷温帯の種類が主」であることが原因のようです。
私が熱を反射・放出するための構造と考えた物を、彼等は熱を吸収しやすい構造と考えたとしたら、それはまったく、逆向きの議論になってしまいます。
しかし、翻って考えてみますと、場合によって、このことはどちらにも適用できることかもしれません。
つまり、多分、冷温帯の高山のガレ場のような場所は地表が黒っぽいでしょうから、反射光のスペクトルは黒っぽいものになる(つまりはほとんど反射されない)と思うのです。
そうすると、大部分の光は吸収されるわけですから、当然、冷温帯の種は、かなりの熱を吸収し、体温を上げることができるのではないかと思います。
逆に、最初紹介したようなテキサスの砂漠など明るくて高温の場所に住む種は、周囲と同じ白〜明るい色を主として反射することになりますから、大部分の熱を放出し、体温を下げる効果が見込めることになると思います。
そういうことを考えると、場所に合わせて、周囲の色に体色をマッチングさせるということが、すなわち、虫自体の熱の吸収・放出にとっても、それぞれに有利に働くことになるのではないでしょうか?
盃刻発色構造の光・熱・風に対する構造的な効果については、今後、物理や材料工学などの専門家の助言や実験が必要でしょう。
専門の方、ご教示をよろしくお願いいたします。
また、推定可能な部分として、高温域では、多層膜自体も幾分なりとも膨張して膜の厚みが増すと思われ、そうすると、反射する光も、いくらかでも青→赤の方向に波長が変化すると思われます。
これは、熱反射が増大する方向ですから、虫にとっても都合が良く、暑い所に進出していく種にとっては、そちらに向かって進化しやすいと思われます。
同様に、低温域では当然、逆の方向へ向かうはずです。
Schultz等の論文の1-1の左上の写真では、前のSchultz(1991)では作画・図示した盃刻発色構造が、きれいな電子顕微鏡写真として、実写されています。
(Schultz等の論文の図1-1-A 左上の写真)
右の図が盃刻1つの断面と思われます。
(Schultz等の論文の1-1-B 右上の写真)
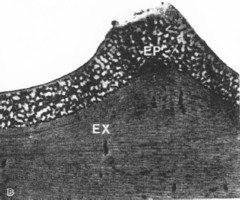
これらの写真から盃刻にも2層(表面下の浅い層と、その下の層)があることは、容易に見て取れます。
この点、私の考えとも一致します。
この写真からは浅い層に多層膜構造は認められず、比較的目の粗いスカスカの層(多分、透明であることが想像される)になっているように見えます。
ということは、この写真からは、Schultz(1991)の言う「盃刻の浅い層に多層膜構造があり、そこで光を反射している」ということは読み取れないように思います。
また、Schultz等は、盃刻発色構造と生態的な機能については詳しく説明していますが、この中央が凹んだ盃様の穴ぼこ(pit)がどうして出来たかという原因や、その部分が元々何のパーツに由来するかということには、言及してはいないようです。
金属光沢と盃刻による色のモザイク配列について、それら互いの形成課程の順番や、どのような意味合いで進化してきたかという点についても、ここでは意識されてはいないようです。
堀さんからも
「Schultzは、上記のように、体色を温度適応と敵からの防衛の2面から捉える立場ですが、盃刻発色構造の機能とその進化傾向などについての言及は見られないようです。
自然科学者として、測定法などを工夫し実証的な研究を目指していますが、このことから進化を直接調べようとは考えていないようです。系統群ごとの盃刻の現れ方についても述べていません。
これについては、私は系統群ごとに盃刻発色構造のあり方を調べるのは大いに価値があると思います。
今坂さんが取り上げた種群以外の多くの種についても調べ、現時点で分かっている系統群ごとの進化傾向などの情報と照らし合わせるなら、さらに面白い研究になると思います。」
との感想をいただきました。
私も、進化は直接調べることができないとしても、何らかの方法で、その一部でも証明しようと工夫することが、進化と多様性をテーマにするもののあるべき姿勢だと考えています。
4回に渡るハンミョウの色の話を、ここまで読んで下さった有意の若い方々に、是非とも、堀さんも期待されているような、さらなる研究を推進して頂きたいと考えています。
<日本産に見られる上翅盃刻発色構造の分化>
最後に、当たっているかどうかは解りませんが、日本産ハンミョウに見られる盃刻発色構造の形成過程(というより、一度完成された盃刻発色構造の、新たな変化の過程ですが)の順番と傾向を、ざっと推定したいと思います。
ただし、この点については今坂単独の見解で、堀さんの承認はいただいていないことを、前もってお断りしておきます。
以上に述べてきたような盃刻発色構造の形成過程を考えると、盃刻が完成された時点で、既に金属光沢を持っていたと考えられます。
特に広義(かつての)のCicindelaなどは、ハンミョウの中でも進化した群と考えられ、世界中に広がっている群であり、ずいぶん以前に既に金属光沢を持つ盃刻発色構造が属レベルで完成されていたと思えます。
しかしながら、日本産の中に何種か、その基本から逸脱しかけた種が見られます。
これらの種に見られる盃刻発色構造の不完全な状態は、アメリカ産で紹介した数種のような、今から発展的に完全な盃刻発色構造が形成される方向と言うより、むしろ、一度完成された盃刻発色構造が新たな適応を模索して、変化(あるいは退化)し始めている状態であるように思えます。
盃刻を持ちながら金属光沢を無くしてしまったホソハンミョウやタテスジハンミョウ、
無くしかけているニワハンミョウ、
グラディーションで色分けして、有る部分は金属光沢を持たず、また有る部分は逆に、目立つ色彩を獲得したハンミョウなどがその例で、
これらの特徴は、むしろ、子孫的と考えて良いのではないかと思います。
これらの種は、地表性ハンミョウの本来の生息場所である、夏のカンカン照りの裸地を生活の場とすることを止めたために、さらに、盃刻発色構造の状態を変化させたのではないかと思われます。
ホソハンミョウとタテスジハンミョウは、生息地を草地の草蔭や林床に移し、ホソハンミョウの方は、ついでにこのような場所での生息に余り必要のない後翅も退化させたのでしょう。
ニワハンミョウは活動時期の主力を春に移し、林縁の木陰を利用することで、金属光沢を無くし、あるいは、無くしかけているのではないかと思います。
そして、ハンミョウの場合もむしろ活動時期の主体を秋〜春にして、暑さ対策をそれほどしなくても良いようにしたのかも知れません。
あの目立つ色は警戒色と思われますが、何に対してでしょう?
毒でも持っていると鳥やヘビ、カエルなどに思わせようとしているのでしょうか?
(堀さんのご教示によると、ハンミョウの色は分断色とする説が有力だそうです。
分断色の説明:
動物の眼は、光と影のコントラストの強い部分があると、そこに焦点が合ってしまい、全体の輪郭がわからなくなってしまう。たとえばシマウマの場合は、捕食者の眼には白い部分と黒い部分とが別々に強調されて映るので、シマウマ全体の輪郭はあいまいとなる。動物は、一つの輪郭つまりシルエットで物体や生物を認識しているので、こうなると、シマウマを一つの生物として認識することがむずかしくなる、つまり見逃してしまうのだ。このように、体の輪郭を分断して見せるカムフラージュ効果のある色を分断色という。)
また、一部の種で増加傾向にある、点刻の前部にある顆粒の意味も、良く解りません。
上翅表面に、さらに、凹凸を増加させて、放熱効果を増大しようとしたのでしょうか?
この部分は、本来は凸〜平坦部分を持ち上げて形成されただけのようで、通常は平坦部と同様の赤銅色の光を放っています。
いくつかの種では、その顆粒の表面に見られた盃刻をも、消滅させる方向に進んでいるようです。
この意味も今のところ不明です。
イカリモンハンミョウとルイスハンミョウでは、さらに、顆粒のみならず凹部以外の全域の盃刻を無くしかけています。
同様にカンカン照りの砂浜に生息する種ですが、何か別の意味か、特別の暑さ対策があるのでしょうか?
以上の考えを纏めると、地表性の盃刻発色構造を持つ日本産ハンミョウ類において、分化(退化)の方向は以下のようなチャートで考えることが出来ます。
金属光沢+盃刻+色のモザイク配列(盃刻発色構造)の獲得
ヨドシロヘリ、エリザ、トウキョウヒメ、ヒメヤツボシ
↓
↓♀に鏡紋→シロヘリ、コハン
↓
↓上翅の白色部分を増加→カワラ
↓
↓金属光沢消失→タテスジ
↓ ↓
↓ 後翅退化→ホソ
↓
表面の凸凹多く、凹部の前に顆粒発達
ミヤマ、コニワ、アイヌ、アマミ
↓
↓後翅退化→マガタマ
↓
↓盃刻発色構造の色のモザイク配列解消
↓ (色はグラディーション)→ハンミョウ、タイワンヤツボシ
↓
顆粒の盃刻を消失傾向
↓
↓顆粒増加→ハラビロ
↓
↓金属光沢を消失傾向→ニワ
↓
凸部全体の盃刻を消失傾向
イカリモン、ルイス
以上は、あるいは全くの今坂の空論に過ぎないかもしれません。
それでも、仮に延々と述べてきた議論の、どの部分かに正鵠を得ている部分があり、これから研究しようという人のヒントになるとすれば、それは望外の幸せです。
最後になりましたが、ハンミョウ類の同定を始め、分類や参考文献の内容についても懇切丁寧に説明していただき、本文についても色々適切な感想と示唆をいただいた堀 道雄京都大学教授に、心よりお礼申し上げます。

