令和元年9月3日に、九州大学名誉教授でゾウムシ類の世界的権威 森本 桂博士が逝去された。
(1997年3月1日退官記念講演中の森本先生:「森本 桂コレクションとゾウムシ学」パンフより引用)

昨年、脳梗塞を発症されて、闘病生活を続けられていたものの、時に車椅子に乗って大学にも姿を見せられ、日本産ゾウムシ類の研究を続けられていたらしい。
日本産ゾウムシ総説3冊目となるキクイゾウムシ科の完成が間近と伺っていたので、特に残念な気がする。先生も心残りであられただろうと思う。
逝去される直前の4月15日から8月30日まで、九州大学総合博物館常設展示室において、「特別展示 森本 桂コレクションとゾウムシ学」の展示が行われていた。


森本先生は1934年高知県生まれ、1952年九州大学農学部に入学。ゾウムシ類の比較形態学を研究。1956年卒業。
“先生の存在を知る”
先生は1961年農林省に就職され、熊本市立田山の林業試験場でマツの害虫とシロアリの分類を研究。松食い虫として有名なマツノマダラカミキリがマツノザイセンチュウの運び屋として、マツ枯れの主要な原因になったという研究結果を発表。
その報告を元に、全国の松林で松食い虫の撲滅のために薬剤の空中散布が行われた。
私と先生の最初の接点は、この空中散布だった。
私は1949年生まれで、先生よりは15歳年下、大学入学後、当時ブームとなっていたカミキリムシの採集に夢中になった。カミキリ幼生期の権威 小島圭三先生に指導していただいて、当時、カミキリの幼生期を解明すべく採集を続けていた。


大学卒業後、実家を継ぐことになり長崎県島原市に帰郷したが、毎日、頭上にヘリコプターが飛び回り、山は立ち入り禁止で、薬剤の空中散布が行われていた。
若くて、ナチュラリストを自認していた私としては、この空中散布は諸悪の根源であるように思われた。そして、地元の島原新聞に、1973年6月20日と21日の2回に分けて「自然は再生できない=島原の農薬空中散布について=」と言う1文を寄稿した。

そして、この空中散布の根拠を与えた研究者が森本先生であることを突き止めたので、この新聞を同封して、空中散布を止めさせていただきたいとの要望の手紙を書いた。
しかし、先生から返事が来ることは無かった。
“先生にゾウムシを見ていただく”
それから5年が経った1978年に、先生は九州大学農学部昆虫学教室に助教授として着任された。
私の方は、帰郷後、地元のカミキリ、そして、甲虫全体のファウナ調査を続けながら、長崎昆虫同好会と長崎県生物学会に入会した。後者によって発刊された「対馬の生物」の好評に勢いを得て、1982年には「五島の生物」が企画されたが、その中では甲虫リスト作成に携わることになった。
特に新種を含めて固有種が多い男女群島のリストも担当したことで、同定等、素人の手には負えず、カミキリは当時昆虫学教室の助手であった槙原寛氏にお願いすることにして、ゾウムシについては森本先生を紹介していただいた。
先生は丁寧に、男女群島産について標本を同定してくださり、 過去の記録についてもご教示いただいた。

この後、未知の新種らしいゾウムシに出会うたびに、先生に連絡を取って見ていただいた。冒頭にある、特別展示のパンフに載っているアカサビシギゾウムシのホロタイプは私が採集した島原産である。

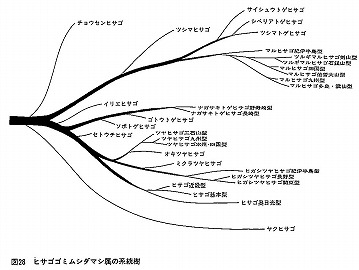
“ヒサゴゴミムシダマシの研究”
この頃また、私は九州大学英彦山実験所所長の中條道崇先生と一緒に、ヒサゴゴミムシダマシ属 Misolampidiusの再検討を進めていた。
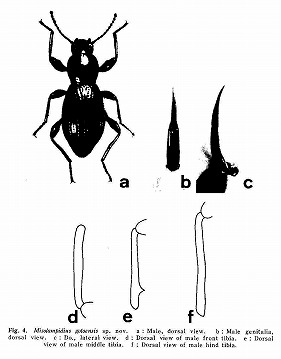
Esakia(19)1982に中條先生と共著で5種を新種記載した後、主として本土産の地域変異を中心に、1983年6月から1984年8月まで、「ヒサゴゴミムシダマシ属の系統と進化」と題して、月刊むしに11回に渡って連載させていただいた。

(ヒサゴゴミムシダマシ属の♂脛節のトゲの進化)
“先生から九大の研究生に来ないかとのお誘いを受ける”
それから10年後の1993年12月5日、長崎大学で日本昆虫学会・日本鱗翅学会の九州支部会の合同大会が開かれた。会には長崎昆虫同好会も協賛して、私も参加した。
大会の一般講演では、楠井善久さんと一緒に、ビロウドコガネ Maladera属の分類と、五島の赤いアオハナムグリについて報告した。
大会後懇親会に移り、森本先生にお酌をしに行ったところ、「九大に来ませんか?」とのお誘いを受けた。
私は英語力に自信が無く、大学卒業後20年近くが経ち、とても入学試験は通らないと危惧したが、先生によると、無試験で、教授枠による社会人の研究生を受け入れる制度があるという。
自分に大学での研究などできるか? まったく自信が無かったが、当時、地元島原市は普賢岳噴火災害がまだ収束しておらず、家業もほぼ立ち行かなくなっており、妻子を抱えて途方に暮れかけていた時期だったので、ともかく、気を奮い立たす意味からも、お話を受けることにした。
まさに、先生のお誘いは、天の救いとも感じられた。
先生に松食い虫のことで手紙を出してから、丁度20年目にあたったが、先生は覚えておられず、あるいは手紙は先生の元に届いていなかったのかもしれない。
“先生にジョウカイボン科の研究の進め方の指導を受ける”
1994年10月4日、この日初めて九大に先生を訪ね、昆虫学教室に研究生として通い始めた。月に1度、先生のご都合を確認した上で、島原から有明フェリーが出る多比良港まで車で走り、荒尾に渡ってから南関で高速に乗り換えて福岡インターで降り、箱崎学舎にある農学部昆虫学教室へ急いだ。
研究テーマはジョウカイボン科(特にクビボソジョウカイ族)の系統分類。その時点で、国内産ジョウカイボン科は120種余りが知られていたが、多分、その倍以上の種が存在すると考えていた。
登校初日、先生はジョウカイボン科の基礎的な文献20編余りのコピーを用意して教授室で待ち構えていらした。その上で、研究を始めるに当たって調べるべき事を列挙したメモと、研究に必要な文献のリストを示され、九大図書館でのそれらの文献の探し方、ズーロジカルレコードの引き方、コピーの取り方などを教えていただいた。
以上の事を教室の他の学生に話したところ、先生にそんなことをしていただいた者はいないと言うことだった。
先生の意気込みも充分に感じられて、1ヶ月ごとにその月に調べたこと、解ったことをメモにまとめて報告した。先生はその上で次回への指針を示され、次になすべき事を教えていただいた。
“相同”
そうした中で最初に強く心に残ったのが「相同」という概念だった。
形態分類学では形を元に分類をする。AとBの形がどこか違えば別種という判断をする。ただ、その違う部位が相同であることが必須条件である。
例えば、まったく形は違うが、鳥の翼とトカゲの前足は相同である。逆に鯨の尾と魚の尾びれは相同ではない。鯨の尾は足の変形したもので、魚の尾は背骨の延長である。つまり形が違っても由来が同じ器官が相同で、似通っていても元は別の器官であったものは相同ではない。
先生は、ジョウカイボン科の分類を進めるに当たって、使用する形質の細かい部分についても、1つずつ、相同関係を踏まえた上での部分名称を付けて、分類を進めるよう指導された。
系統分類に必要な最も重要な形質として、雄交尾器と、頭部の内部骨格であるテントリウム(Tentorium: 膜状骨)を調べてみることを勧められた。
“雄交尾器”
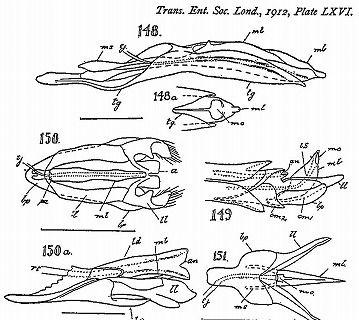
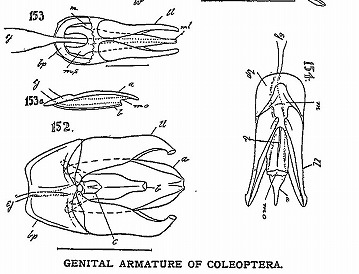
雄交尾器の教科書としては、100年以上前に出版されたSharp・Muir(1912)を勧められた。
Sharp, D. & F. Muir, 1912. The comparative anatomy of the male genital tube in Coleoptera. Trans. Ent. Soc. London part 3: 477-642. (Reprinted by Ent. Soc. America, 1969).
この本には甲虫のほとんどの科の雄交尾器が図示、解説され、全ての部位に部分名称が付けられている。
これらの図を分類群ごとに眺めると、科あるいは上科ごとにほぼ同じ構造をしているものの、系統的に離れた群では、まったく違った構造や形をしている。
(Sharp・Muir, 1912のpl. 56 三片型など: No. 149-150ツツシンクイ科、No. 151ナガハナノミ科、No. 151クシヒゲムシ科、No. 154コメツキムシ科)
先生は彼らの論文も含めた集大成として、原色日本甲虫図鑑(I)のp63に雄交尾器の模式図を書かれている。ここでは甲虫の雄交尾器を7つに類型化されている。
(甲虫の♂交尾器の7つのタイプ: 森本, 1984より引用)
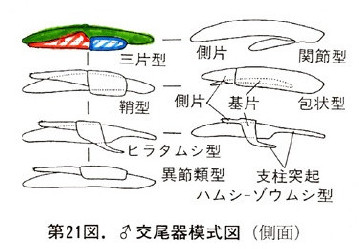
甲虫の最も基本的な形として、上記の三片型が相当すると思われるので、以下に示してみたい。
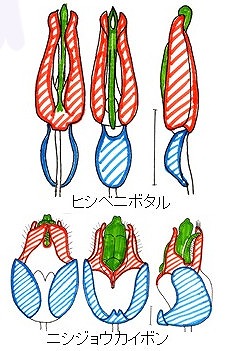
このヒシベニボタルの♂交尾器が典型的な三片型(左から背面、腹面、側面)であり、青色部分が基片、中央の緑部分が中央片、赤色の部分が側片である。同様に、下段のニシジョウカイボンについても相同部分を同色で示している。
甲虫では図鑑の前1/4付近に置かれているミズスマシ、ガムシ、シデムシからコガネ、ナガハナノミ、タマムシ、コメツキ、ベニボタル、ジョウカイボン、ホタルなど、主として幼虫が水棲のグループでこの様な三片型の♂交尾器が見られる。
典型的な三片型としたヒシベニボタルでは、♂♀がお互い後ろ向きにお尻同士をくっつけ合う形で交尾を行い、♂交尾器が♀腹部に挿入される。これを逆向き型交尾姿勢と呼ぶ。
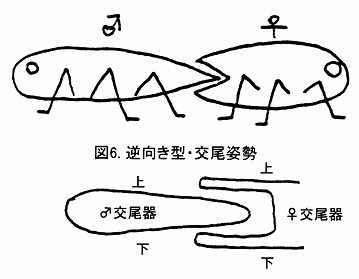
当然ながら、雌雄の♂交尾器はお互いの上下関係はそのまま同じ位置で交接する。
甲虫以外では、カメムシ目がこのタイプであり、他にも背面から腹節を横回しにして挿入するバッタ目や、腹合わせに抱き合うような形で交尾するチョウ目なども、雌雄の交尾器の上下関係が一致しているという点で、同じ交尾姿勢の発展形と考えることが出来る。
ジョウカイボン科では、Malthodini族において、逆向き型交尾姿勢が見られ、雄は種ごとに異なった複雑な形の腹板と背板との間に雌の腹節を上下から挟み込んで、♂交尾器を挿入する。
ジョウカイボン亜科においても、ニシジョウカイの例を示したように、構造的には三片型である。
(ニシジョウカイボンの♂交尾器)
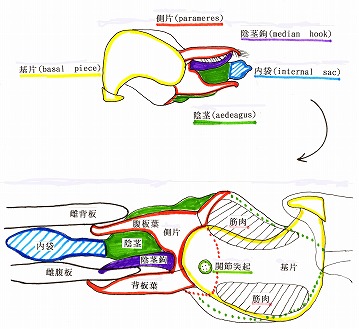
しかしながら、ジョウカイボン亜科の♂交尾器は、Malthodini族とは異なり、交尾姿勢は♂が♀の背側に位置して交尾する馬乗り型で、♂交尾器は上下逆になって♀交尾器に挿入する。
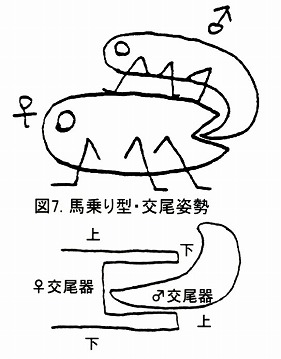
この時、側片の一部分である背板葉(Dorsal plate)と中央片の背面に生えた陰茎鈎(Median hook)によって、雌の腹版を挟み込んで把握しながら、♂交尾器の先端部を雌体内に挿入するというのが、ジョウカイボン亜科の特徴である。
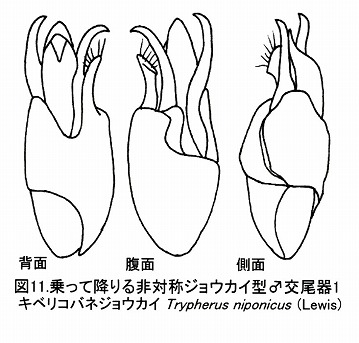
さらに、コバネジョウカイ亜科では、基片、中央片、側片共に、時計回りに先端がねじれている。
(コバネジョウカイ亜科の♂交尾器 乗って降りる非対称ジョウカイ型:キベリコバネジョウカイ)
このタイプでは、以下の模式図のように、雄が雌の背面に位置して挿入を開始し、雌の右へ降り、さらに反対側を向くことで、♂交尾器は雌の体内で半回転しながら挿入されることになる。
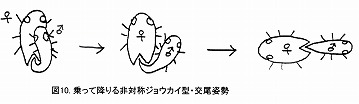
コガネムシの一部や、ベニボタルの一部など、左右不対称の♂交尾器を持つ種は、この様な形で挿入すると思われる。
つまり、構造的には三片型の♂交尾器を持つジョウカイボン科内に、実際の交尾姿勢には逆向き型(Malthodini族)、馬乗り型(Malthinini族、ジョウカイボン亜科、クシヒゲジョウカイ亜科)、乗って降りる非対称ジョウカイ型(コバネジョウカイ亜科)の3つの交尾姿勢のタイプがある。
Sharp・Muir(1912)の図版を見たり、自身で、手持ちの様々な科の♂交尾器を摘出して見てみたりしても、殆どの科では、おおよそ1つの♂交尾器の型だけで、交尾姿勢も馬乗り型1型だけに限定されていた。
このことから、1つの科の中に3つの交尾姿勢のタイプが存在するジョウカイボン科は、甲虫の中でもかなり特殊な(原始的な)科であるとの感触を得た。
“Tentorium 膜状骨”
次に、膜状骨である。こちらは、昆虫の頭部にある内骨格で、その機能として、頭蓋内で頭蓋の強度を保つと共に、食道などの消化器系、目、触覚、大あご等の神経系、大あご・触覚等を動かす筋肉系の3つの要素を分断配置し、交通整理を行っていると考えられる。
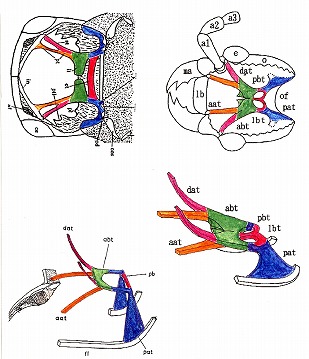
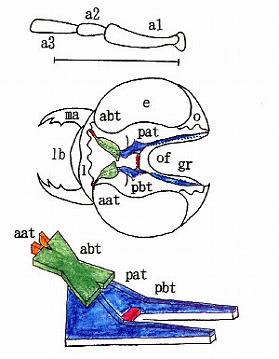
以下の図に於いて、左上はStickney(1923)による膜状骨の背面からの模式図(Fig. 443)。彼は甲虫のほぼ全てのグル-プに渡って膜状骨を図示し、特徴などを詳細に記述した。
さらに左下はEvans(1962)による膜状骨の構造を立体的に表現した模式図(Fig. 3)である。右上は私が書いたオオモモブトシデムシの背面から見た膜状骨で、右下はEvans(1962)をまねてそれを立体模式図にしたもの。全ての彩色は、相同部分について私が行った。
Stickney, F. S., 1923. The Head capsule of Coleoptera. Illinois Biol. Monogr. 8., 104pp.
Evans, M.E.G., 1962. The muscular and reproductive systems of Atomaria ruficornis (Marsham) (Col., Cryptophagidae). Trans. Roy. Soc. Edin., 64(14): 297-399.
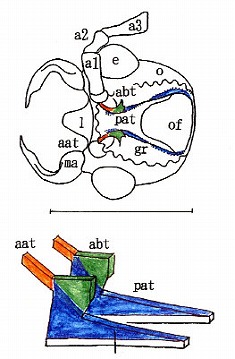
甲虫の最も基本的(原始的)な構造としては、左右の立ち上がった柱(pat)の間に2本の橋梁(緑部分であるabtと、赤色部分であるpbt)が存在することである。
(基本的な構造を持つハネカクシ、カツオブシムシの膜状骨)
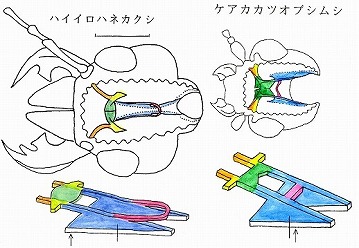
abtとpbtは図鑑の最初にあるハンミョウなどから、最後近くのカミキリまで、まんべんなく見られ、膜状骨においては、♂交尾器のような科や上科などのグループによる固有性はあまり見られない。
しかしゾウムシ類では左右の立ち上がった柱(pat)部分が癒着したり、abtを欠いたりと、かなり退化(子孫)的である。
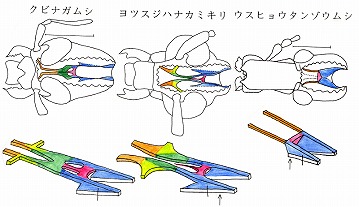
ゾウムシ類のように、グループによっては1つの科の中で、亜科、あるいは族レベルで、構成するパーツのある部分を欠くグループがあり、その科の中で進化的(子孫的)な形質と見なすことが出来る。先生はゾウムシの例から、ジョウカイボン科でも膜状骨を調べてみることを勧められたのかもしれない。
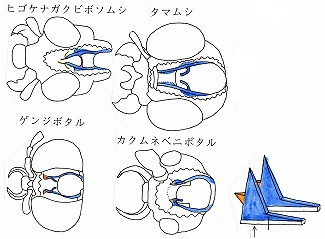
以上に示した例のように、例えばゲンジボタルのように複眼が特に拡大したり、あるいは、大顎を動かす筋肉が拡大したりして、頭部内部のこれらの容量が拡大した種などでは、膜状骨の一部(橋梁部分のabt、pbtなどが欠ける事が多い)、あるいは大部分が退化する傾向がある。
ジョウカイボン科では、この膜状骨の構造が、亜科ごとに異なっており、亜科を分ける良い形質となった。
ジョウカイボン科の4つの亜科の中で、abtとpbtの基本構造を僅かながら残しているのは、コバネジョウカイ亜科のみである。
(チビジョウカイ亜科の膜状骨、左:Malthinini族、右:Malthodini族)
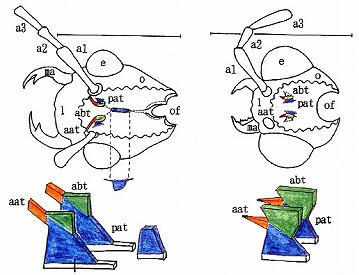
チビジョウカイ亜科はかなり退化(子孫)的で、特に、Malthodini族では支柱部分のpat以外がほぼ消失している。
(ジョウカイボン亜科の膜状骨、左:ジョウカイボン族、右:クビボソジョウカイ族)
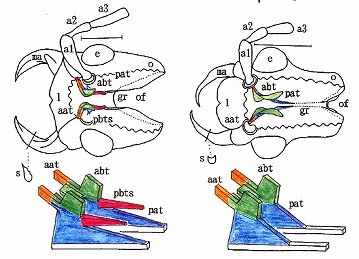
ジョウカイボン亜科では、やや退化が少なく、ジョウカイボン族はpbtの名残と言えるトゲ状突起(pbts)がある分、クビボソジョウカイ族よりは祖先的である。
クシヒゲジョウカイ亜科はほぼジョウカイボン亜科と同じ。
膜状骨の探索には、大部分の甲虫のみならず、ゴキブリ、ハサミムシなど他の目までおよび、先生にこれらの目でも基本的には甲虫と構造が同じ事を報告した。
(ゴキブリ、ハサミムシなどの膜状骨)
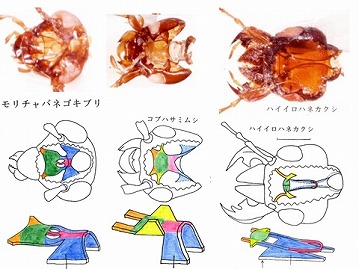
しかし、先生は「いいかげんにしろ」と怒られ、ジョウカイボン科の議論に、類縁関係が遠すぎるゴキブリ、ハサミムシなどを持ち出すのは無意味だと断じられた。
確かに、膜状骨の探索に興をそそられて、自身が何の目的でそれをやりだしたのか、スッカリ忘れていた。
“転職”
話は少し遡る。九大に通い始めて半年ほど経った頃、いよいよ家業の先行きが怪しくなったので、丁度、福岡市に支店を開設すると言うアセスメント調査会社に勤務することにして、通勤可能な久留米市に引っ越すことになった。これで大学も距離的・心理的に相当近くなった。
先生はその話を聞かれて、半分研究職でもあり、喜んで励ましてくださった。それでも、「あまり仕事をし過ぎないように・・・」とのアドバイスもいただいた。
会社の方には1995年9月から出社したが、月1回の登校も許可いただき、遠隔地での学会等への出席も年2回程度は配慮いただいたので、研究環境としては、島原時代より向上した。
“系統解析”
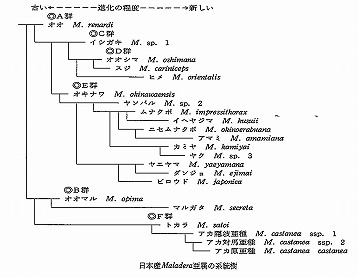
♂交尾器と膜状骨の検討に加えて、大顎、上唇、頭盾、咽喉縫合線、頭部の傾き、触角、上翅、爪など、それぞれの形質について、どのような形が祖先的であり、それがどう変化して子孫的な形になったかということを念頭に置きながら、形質評価を進めていった。
クビボソジョウカイ族がジョウカイボン科の中で、どのような位置にあり、さらにホタル上科内で、また、甲虫全体の中での位置を確認するのが目的であった。
先生はその目的のために、なるべく有意な多くの形質を選定することを指導され、結局48項目の形質を抽出して表を作り、系統解析ソフトにかけて分岐図を求めた。
その結果、ホタル上科の中では、ジョウカイボン科はホタルモドキ科に一番近く、ほぼCrowson(1972)が示した分岐図と同じであった。
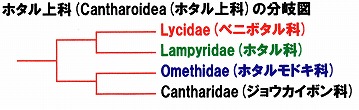
(Crowson, 1972による分岐図)
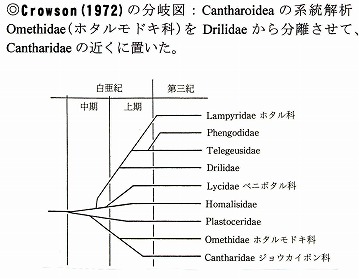
Crowson, R. A. (1972) A review of the classifification of Cantharoidea (Coleoptera), with definition of two new families, Cneoglossidae and Omethidae. Revista Universidad Madrid, 21(82): 35-77.
また、ジョウカイボン科の中では、クビボソジョウカイ族はジョウカイボン族と共にクシヒゲジョウカイ亜科に近いことが明らかになった。
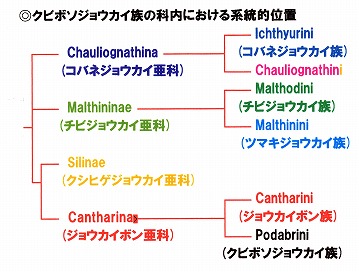
世界的なジョウカイボン科の権威Wittmerの一番弟子と目されるBrancucci(1980)は、自身の系統解析に基づいて以下のような分岐図を発表している。
(Brancucci, 1980の分岐図を見やすいように改変)
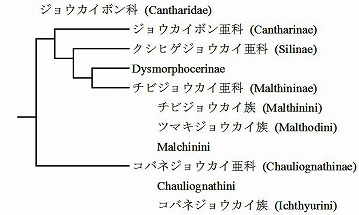
Brancucci, M., 1980. Morphologie compar?e, ?volution et syst?matiqe des Cantharidae (Ins.: Col.). Ent. Basil., 5: 215-388.
原色日本甲虫図鑑(III)などのジョウカイボン科の配列は、概ねこの考えに基づいていると判断される。科のトップにクビボソジョウカイ族がきて、次いでジョウカイボン族、チビジョウカイ亜科、クシヒゲジョウカイ亜科、コバネジョウカイ亜科の順である。
佐藤正隆, 1985. ジョウカイボン科 (in 黒澤良彦ほか, 原色日本甲虫図鑑 III), :107-118, 保育社.
私の分岐図との一番大きな違いは、Brancucciはクシヒゲジョウカイ亜科とチビジョウカイ亜科が近いとしている点で、ジョウカイボン亜科はその外側に来ている。
私のはむしろ、ジョウカイボン亜科とクシヒゲジョウカイ亜科が近くて、チビジョウカイ亜科とコバネジョウカイ亜科は、その外側に、並列に付く。
また、系統解析の中で、祖先的、子孫的を形質ごとに判断した結果、コバネジョウカイ亜科が最も祖先的で、チビジョウカイ亜科がそれに続き、クシヒゲジョウカイ亜科、ジョウカイボン亜科が最も子孫的との結論に達した。
さらに、ジョウカイボン亜科の中では、ジョウカイボン族がより祖先的で、クビボソジョウカイ族がより子孫的という結果も得た。
つまり、上記図鑑の配列はほぼ逆転することになる。
“旧Podabrus属の分割”
さらに、クビボソジョウカイ族の中においても、従来、Podabrus属に含められていたものの内、Podabrus、Hatchiana、Malthacus、Asiopodabrusの4属に分けることが出来るとの結論に達した。
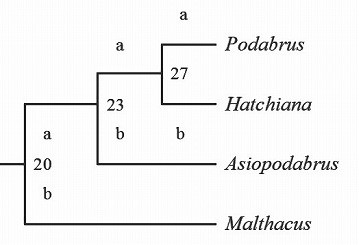
こうして、分けてみると、Podabrusは新大陸(北アメリカ)が分布の中心で、100種近い種が分布し繁栄していて、旧大陸の北部(ヨーロッパから北海道)にも少数の種が分布する。
Hatchianaは新大陸に数種と日本を含むアジア東岸の中緯度地域に10数種程度が分布し、どちらかというと東アジアが分布の中心、Malthacusは周北極海と呼べるほど高緯度地域(北海道を含む)のみに10数種が分布する。
そして、Asiopodabrusは国内で100種以上分布するにもかかわらず、それ以外では、台湾、東アジア地域に数種が知られるだけで、正に、日本列島が分布の中心と言える。
属ごとに繁栄している分布の中心が明らかに異なり、由来の違う群(別属)で有ることを確信した。
“種と亜種”
属レベル以上の検討がひとまず終了したので、次に種レベルの検討に入った。
ジョウカイボン科の中でも、特にAsiopodabrus属の地域性と地域変異は凄まじく、地域ごとに微妙に変異していく個体群について、何処までが原種で、どこからが亜種になり、そして、どこから別種と判断すべきか、分類学としての基準はないものかと思い、先生に確認してみた。
先生は「そんなものは無い」と答えられた。「研究者が文献や標本を繰り返し研究して、これは種、これは亜種と、確信できる目が形成された時、それが種であるし、亜種である」との禅問答のような答えであった。
数多くの研究者が個別に見解を発表することによって、自ずからコンセンサスが取れ、定説として確定していくものらしい。
実際には、種と亜種の割り切り方について、研究者によって好みもあるようで、先生は、初期の研究を除いて、基本的に亜種は作られず、形態的に違うものは種と判断されているようである。
あるいは、扱う分類群の性格にもよるかもしれない。
ジョウカイボンは、とても、種だけでは割り切れない。亜種と、できれば、さらに下のランクの分類単位が必要な気がする。
少なくとも、先生に対して、そう断言できるだけの目は養えたと言うことだろう。
“先生の退官”
森本先生は1997年3月に退官された。しかし、退官後も昆虫学教室の標本室の中に研究場所を確保されて、平日は毎日出勤して午前中は研究を続けられていた。
私の研究は道半ばで、後任の教授からは、引き続き森本先生の指導を受けるように告げられた。
先生も、退官時にはまだ私の研究が完成しないことは最初から織り込み済みで、引き続き、毎月標本室に出向いて指導していただいた。
先生の退官前も、退官後も、教室の学生の話では、「今坂さんほど、先生とコンスタントに話をしている者はいない」ということで、よほど先生にとっては、気がかりな生徒であった模様である。
“論文の執筆”
旧Podabrus属内の分類がほぼ確定したので、まず、Hatchianaの日本産の総説を、先生の指導の下で行った。記載文は全面的に先生に添削していただき、あまり得意でない描画も、先生の示唆で、できるだけ大きく描いて、グッと縮小することでなんとか仕上げた。
Imasaka, S., 2001. Taxonomic study of the genus Hatchiana in Japan (Coleoptera, Cantharidae, Podabrini). Jpn. J. syst. Ent., 7(2): 279-313.
この論文が公表された後、別刷りを持参したところ、「スッキリまとまった論文になりましたね」と褒めていただいた。
(Imasaka, 2001の付図)
しかし、先生からは、その後、日本に1種しか分布しないPodabrus属や、2-3種のMalthacus属は別にして、70種以上分布するAsiopodabrus属についても、種レベルでの検討をするよう指導された。
一方で、この時期までに、私とは別にAsiopodabrus属の研究を続けられている高橋和弘氏と桐山功氏が、断片的であったが盛んに多数の種の記載を発表され続けていて、既知種だけでも100種を超えていた。
“大学と会社をやめる”
一方で、私自身、本業のアセスの仕事が本格的に忙しくなり、月1の登校も難しくなってきた。また、研究生での在籍も7年半に及び、教授からも論文の期限を切られてしまった。
と言うことで、とうとう、2002年の春、論文を提出することなく研究生を辞退した。
先生からは、「論文を提出さえすればD論として受領できたのに・・・」とグチられたが、時間的にも、能力的にもまったく無理と言わざるを得なかった。
結局、私は、先生のご期待にまったく添うことが出来ない不詳の弟子となったわけである。当初の先生のご厚意と熱意を知るだけに、極めて心苦しかった。
さらに、そのアセス会社の方も、あまりの忙しさと会社内での自身の扱われ方に不満が生じ、2004年3月に退職。その後、E-アシストとして独立して、アセスメント調査の現地調査と同定業務を始めた。
“「日本産のアオハムシダマシ属のまとめ」が原因で破門に?”
退職後、ジョウカイボンとは別に、少し前から調べていた日本産のアオハムシダマシ属のまとめがほぼ出来上がったので、先生に、記載部分などの英文の添削をお願いしたところ、快く引き受けていただいた。
比和科学博物館の中村慎吾氏のご好意で、多数のカラーページと本文ページを使用させていただいたアオハムシダマシ属のまとめが出来上がったのは、2005年3月である。
今坂正一, 2005. 日本産アオハムシダマシ属の再検討. 比和科学博物館研究報告, (44): 61-162.
(日本産アオハムシダマシ属の再検討 付図:ホロタイプなどの写真)


さっそく先生に届けたところ、意に反して、先生はたいそうお怒りということであった。
この後の研究者の利便を考えて、Lewisや益本氏等の原記載論文をそのままコピーして本文中に載せたので、その事は盗作に当たるのではないかとの話しも出たそうである。
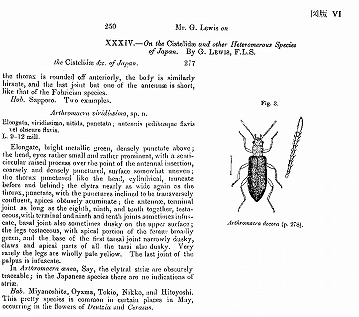
しかし、ちゃんと原典も明記し、そこから引用したことも書いているので、私は研究論文としては、盗作とか、著作権の侵害には当たらないと判断して掲載した。
それでも、原図1つであっても、必ず著者の了解を取って引用されていた、かつての研究者の常識からは外れていて、顰蹙を買ったものらしい。
併せて、先生は、「論文は必要最小限の表現で、読者に確実に解るように書くべきである」と話され、それを実践しておられた。
しかし、私のアオハムシダマシには、分類に必要なこと以外の、調べ始めた動機とか、調べていく経緯とか、アオハムシダマシに関してさまざまに考えたことなどを日本語で縷々書き連ねていて、むしろ、こちらを書きたいが為の論文だった。
先生に添削をお願いした英文にはこれらは含まれておらず、先生はこの英文とその要旨の日本文のみと考えられていたわけで、思いもしないそれに倍する、先生から見れば不必要な日本文が延々と続いていたわけである。
その後しばらく先生には連絡できず、あるいは、「破門?」になったような気分でいた。
“真の原因”
後に、ある人から伺った話によると、先生のお怒りと落胆は、そういうことより、私の矯正(分類学者として正しい道に導くこと)ができなかったことにあったらしい。
想像するに、この文の始めの方に書いたヒサゴゴミムシダマシの連載(大学に行く前)と、アオハムシダマシのまとめ(大学に行った後)と、先生の意図とは裏腹に、まったく変わっていないと判断されたものと思われる。
それだけ、先生が私を買いかぶってくださっていたかと思えば面はゆい気がするし、実際は、私の能力と性質を見誤まれていたのだと思うほかない。
どちらにしても、返す返す、先生にとっては無駄な時間を費やしていただいたわけで、心からお詫び申し上げたい。
“先生に感謝”
先生からはしばらく連絡が無かった。
それでも、しばらくぶりに学会でお会いした際、計画中の日本の昆虫3巻のゾウムシ科(1)で解説されるヒゲボソゾウムシ属の地域変異、色彩変異について、「ゾウムシも、まるで、ジョウカイボンみたいになってきましたよ」と感慨深げに話されたのを覚えている。
また、その後、北隆館の原色昆虫大図鑑IIの改訂でジョウカイボンの部分を担当するようにとの指示があったし、2012年には昆虫と自然の「甲虫類の新しい採集法」の特集で、ハムシやゾウムシなどの採集法として何か書くように、とのお便りをいただいた。
その頃嵌まっていた「草掃き採集のすすめ」という採集方法の報告を書いたところ、「今、まさに目の前で採集しているようで、情景が浮かび、なかなか楽しい報告でした」との評をいただいた。
(草掃き採集のすすめ:タイトルとカラー図版)

最後にお会いしたのは、2015年11月に北九州の「いのちの旅博物館」で開催された日本甲虫学会の大会であった。
だいぶ歳を重ねられていたが、退官後20年近くになるものの、それでも毎日大学へ出勤して研究を続けられていると言うことだった。
日本の昆虫関係の研究者で、勤務や仕事と関係なく、これほど情熱と使命感を持って永年研究を続けられた方はいらっしゃらないのではないか、そう思った。
そんな先生に指導していただいたおかげで、少なくとも虫屋としての現在の私は存在しているわけで、勘違いにしろ、先生が私を誘っていただいたことについて心から感謝している。
森本先生、長い間のご研究ご苦労様でした。後は弟子のどなたかがちゃんと片付けられると思います。旅立たれた後、安らかにお休みください。 合掌