
さて、延々と、島原半島と多良山系のファウナ比較について述べてきましたが、そろそろ、本題に入りたいと思います。
ファウナ全体としての比較では、その2で述べた以上の細かい内容を知ることが出来ません。
当然、地史や分化時期などについての考察の材料にもなりません。
そのため、この後、固有種や特徴的な分布、変異などを示す種について、細かく検討してみたいと思います。
<サンカクスジコガネ九州亜種 Anomala triangularis kyushuensis Nomura>
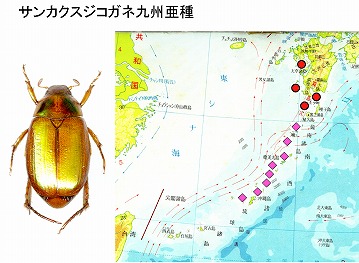
この種は、島原半島南端の口之津町と、東端の島原市で見つかりました。他に、佐多岬と甑島、黒島などの分布が知られ、原亜種がトカラ以南〜沖縄本島に分布しています。九州亜種は原亜種と区別できないとの意見もありますが、今のところ、シノニムには成っていないようです。
本種以外にも、オキナワコアオハナムグリ、シマトゲバカミキリ、ミスジケシタマムシ、セアカケブカサルハムシ、ムモンチャイロホソバネカミキリなど、より、南の方に分布する種が何種かこの地域で見つかります。
これらは、対馬海流沿いに主として九州西岸に多く見られ、大分県など東岸では見つかっていません。海流分布で有れば、四国の南端に沿って親潮が流れていますので、東岸でも見つかってよさそうなのに、西岸だけに分布している理由は、今のところ不明です。
初島・新(1956)は、植物について、このような分布を注目し、「九州西廻り分布」の名の下に、ナタオレノキ、アオモジ、リュウキュウエノキなど25種を紹介しています。
私もその考え方を甲虫に拡げて、今坂(1979)として紹介しました。
・初島住彦・新 敏夫(1956) 九州西海岸に特殊な分布をする植物について. 植物分類地理, 16(4): 98-100.
・今坂正一(1979) 九州西廻り分布をする甲虫について. 長崎県生物学会誌 (17): 5-16.
<ヨドシロヘリハンミョウ Cicindela inspecularis W.Horn>
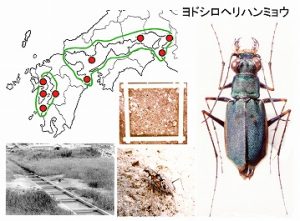
当時、島原半島南端の口之津町在住で、東京農大出身の布施さんから、シロヘリハンミョウの不明種を手渡され、友人を通じて、ハンミョウの生態研究の第一人者京都大学の堀さんに届けたところ、ちょっと変わったヨドシロヘリハンミョウと判断されて、島原に調査に見えたのが、1981年8月でした。
この頃、ヨドシロヘリハンミョウの分布は、大阪淀川河口から大分県臼杵市までの瀬戸内海沿岸と考えられており、島原は飛び離れた産地で、形態的にも、微妙な差が認められました。
私と堀さんの探索で、当日すぐに口之津町の生息地が見つかりました。そこは、ヨシ原と水辺の隙間のごく微妙な空間で、多数の巣穴が密生しており、生息地はジャリ置き場のワキに有るために、人為的に即座に破壊されてもおかしくない状況であることが判明しました。それで、急ぎ「シマバラシロヘリハンミョウ(仮称)の発見と保護について」と題して、報告しました(今坂・堀, 1982)。
後に、堀さんは、大陸産と島原産も、ヨドシロヘリハンミョウに含めています。
分布図には、現在までに文献等で確認した本種の産地を、赤丸で示しています。
有明海特産の魚類などの分布要因を説明した文献を見ますと、氷期の海面が低下した時期には、揚子江の河口が大陸棚を経て九州中部あたりまで達していたようです。その先には、古瀬戸内海が、九州を二分して琵琶湖当たりまで繋がっている時期もあったようです。
その時期に、大陸から、有明海〜瀬戸内海〜琵琶湖と、海岸沿いに淡水性の種や、あるいは、内海性の種が分布を拡げたように記述してあります。
本種の分布もまさしく、その様な形になっていまして、古瀬戸内海とのかかわりが想像されます。
・今坂正一・堀 道雄(1982) シマバラシロヘリハンミョウ(仮称)の発見と保護について. 長崎県生物学会誌 (23): 7-11.
<アリアケホソヒメアリモドキ Leptaleus sasajii Sakai et Telnov>
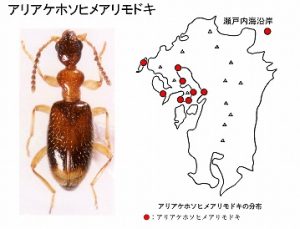
本種は、今坂(1979)において、種名未決定のまま、ホソアリモドキの一種として図示したのが最初の記録です。
体長3mm前後、名前のようにアリに似た褐色の甲虫で、前胸背は赤褐色、上翅の後半に黒褐色の横紋を持ち、♂触角の第5〜6節が三角形に広がるのが特徴です。
Sakai・Telnov(2001)により、国内で初のLeptaleus属の種として新種記載されましたが、当時は、熊本・佐賀・長崎県に分布し、有明海と九州北西部沿岸地域の海岸に限って生息する固有種と考えていました。
最近、平野さんから、瀬戸内海でも採れたとの私信をいただき、比較標本を送って、本種と確認されました。
このような分布から判断しますと、ヨドシロヘリハンミョウとほとんど、同じような分布域を持つのではないかと想像されます。
砂礫海岸や砂浜、河口のゴミの下、草の根際などでみつかりますが、生息地は汽水域に限定されるようで、海岸でも砂浜から2〜30m以上離れた後背地の草地ではみつかりません。今のところ、それ以外の生態は未知で、興味深い種類です。
・今坂正一(1979) 九州西廻り分布をする甲虫について. 長崎県生物学会誌 (17): 5-16.
・Sakai, M. & D. Telnov(2001) Leptaleus sasajii, a new species and first representative of the genus Leptaleus from Japan (Coleoptera, Cerambycidae). SUKUNAHIKONA, Spec. Publ. Japan Coleopt. Soc. Osaka, (1): 327-332.
<タラメツブテントウ Sticholotis sp.>
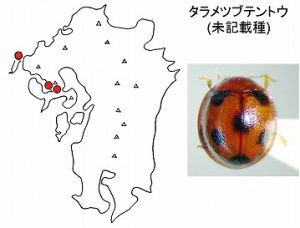
体長2mm前後で、赤褐色、上翅には6黒紋があります。長崎県(大村市黒木、高来町轟の滝)の記録があり、昨年、平戸の安満岳でも採れています。
多良山系の低山帯(標高200〜300m付近)では、常緑広葉樹林内の林縁の葉上や樹皮下などで見つかります。
昨年、タブの樹幹の幹掃き採集によっても、本種が得られることを報告しました。
当初はメツブテントウ Sticholotis substriata Crotchとして記録しましたが、より小型で表面の点刻が密で大きく、斑紋パターンはむしろ、ムツボシテントウ Sticholotis punctata Crotchに似ていますが、黒紋はより小さいことにより区別されます。
未記載種のようで、他の地域からは報告されていません。
・今坂正一(2007) 2001年以降に長崎県多良山系で採集した甲虫-幹掃き採集など新しい採集法の紹介をかねて-. こがねむし, (72): 7-17.
<ヒメチャイロコガネ>
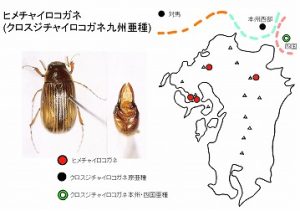
体長10mm前後、全体褐色でツヤのあるコガネムシで、脊振山系と多良山系の低山〜山頂付近に分布し、広葉樹林内の花上や、コケの下より得られています。最近、九重から記録され、西九州の固有種ではないことが明らかになりました。
また、従来は、独立種として扱われていましたが、最近、小林・藤岡(2008)により、近隣の朝鮮半島と対馬、本州と四国のそれぞれ一部に分布するクロスジチャイロコガネ Sericania fuscolineata fuscolineata Motschulskyの亜種に降格され、クロスジチャイロコガネ九州亜種 Sericania fuscolineata minuscula Nomuraとして扱われています。
なお、さらに、本州と四国の大部分の地域には亜種ssp. nipponensis Nomuraが、
北海道には亜種ssp. ezoensis Nomuraが分布しています。
・小林裕和・藤岡昌介(2008) 日本産チャイロコガネ属(コフキコガネ亜科ビロウドコガネ族)について. Kogane, Tokyo, Suppl. 2: 35-76.
<キュウシュウクビボソジョウカイ Hatchiana kyushuensis (Takakura)>
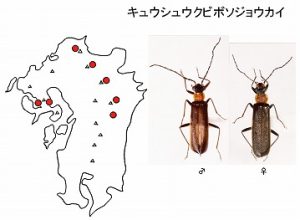
体長10mm前後、頭と上翅、肢が黒色、前胸背が赤褐色になるジョウカイボンで、故・高倉康男さんが晩年に自ら新種記載したジョウカイボン4種2亜種・ジョウカイモドキ1種のうちの1種です。
九州北・中部の英彦山・九重山・祖母山・大崩山の各山系と多良山系の山地帯(標高300m以上)で記録されています。
この後述べるタラクビボソジョウカイに似ていますが、肢が黒いことと、♂交尾器の違いなどで区別されます。広葉樹林内の林縁で、落葉広葉樹の花上・葉上などで見つかります。
・高倉康男(1988)九州産Podabrus属の1新種. 北九州の昆蟲, 35(3): 152.
<コナガキマワリ(オオヒョウタンキマワリ) Eucrossoscelis araneiformis (Allard)>
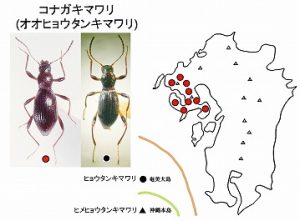
体長7mm前後、弱い銅色光沢を持つ細いヒョウタン型のゴミムシダマシで、後翅は退化し、ちょうど、ヒサゴコミムシダマシを小型にして細長くしたような体形をしています。
本種の分布は、長崎・佐賀両県に局限され、伊万里市と諫早市を結ぶ線の西側(国見山系、黒髪山、西彼杵半島、多良山系、長崎半島と島原半島)のみで記録されています。
平地から山地まで標高にかかわらず広葉樹林の立ち枯れ、枯れ枝などに集まりますが、成虫は、入梅前後の短期間のみに見られ、その時期を逃すと採集できません。
本種は1894年にG. Lewisが自らの採集品をStrongylium helopioides Lewisとして新種記載した種ですが、その後国内からは再発見されないままで、中根(1972)は幻のゴミムシダマシとして解説しています。
今坂(1977)はその幻の種を約80年ぶりに国内から再発見したもので、今坂(1979)でその分布の概要を解説しました。
その後、中條ほか(1983)は上記学名、およびオオヒョウタンキマワリの和名を提唱しています。
同属の近縁種ヒョウタンキマワリ E. broscosomoides Nakaneが奄美大島に、ヒメヒョウタンキマワリ E. michioi M.T.Chujoが沖縄本島に分布しており、本種とは色彩や♂交尾器の形で区別されます。
・中條道崇ほか(1983)ゴミムシダマシ科. 原色日本甲虫図鑑, (?): 341, 保育社.
・今坂正一(1977)コナガキマワリの記録. Elytra, 5(1): 18.
・今坂正一(1979)九州西廻り分布をする甲虫について. 長崎県生物学会誌, (17): 5-16.
・中根猛彦(1972)欧米の自然史博物館を訪ねて(2). 昆虫と自然, 7(7): 18.
<ナガサキトゲヒサゴゴミムシダマシ Misolampidius clavicrus (Marseul)>
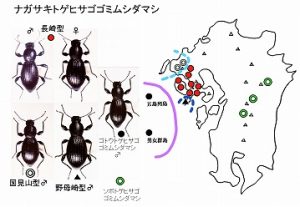
体長15mm前後、ヒョウタン型で漆黒のゴミムシダマシで、後翅は退化しています。コナガキマワリとほぼ同じ伊万里市と諫早市を結ぶ線の西側(国見山系、黒髪山、西彼杵半島、多良山系西部、長崎半島)に分布し、本種は、島原半島には分布していません。
平地〜低山地(標高500m以下)の広葉樹林や、隣接するマツ林、スギ植林内の立ち枯れ、樹皮下などでみられます。
背面の点刻が大きく、雄の中脛節の先端前内側に鋭いトゲを持つことによりMisolampidius属の他の種と、簡単に区別されます。
多良山系東側や島原半島に分布するツヤヒサゴゴミムシダマシとは、ごく近い距離で棲み分けていまして、今のところ、同時に採集されたことはありません。
本種には背面の点刻の状態、♂交尾器などに地域変異があり、
国見山型(図左下)が佐世保市背後の国見山地に、
基本型(図上)が西彼杵半島・多良山系・長崎市周辺に、
野母崎型(図中下)が野母半島先端部に分布します(今坂・中條1983)。
これらの型は亜種程度の差が認められますが、亜種としての記載は行われていません。
五島には本種に近縁なゴトウトゲヒサゴゴミムシダマシ Misolampidius gotoensis M.T.Chujo et Imasakaが分布しますが、後者は体長と体表の点刻が小さく、雄脛節のトゲの形や、♂交尾器の形により区別されます。
本種はG. Lewisの採集品を基にMarseul (1876)によって記載された種ですが、戦後長い間、この学名は国内各地に広い分布域を持つツヤヒサゴゴミムシダマシにあてられていました。
中條・今坂(1982)は、長崎・佐賀両県に分布が限定される本種が、真のMisolampidius clavicrus (Marseul) であることを指摘し、ツヤヒサゴゴミムシダマシには別途Misolampidius okumurai Nakaneの学名をあてるよう提唱しました。
♂交尾器やトゲの形状などから、ゴトウトゲヒサゴゴミムシダマシが本種の祖先形と考えられ、五島と本土とが陸続きになった頃に佐世保付近から本土に侵入し、その後本種に分化し、さらに、3個体群に分化したと考えています。
・中條道崇・今坂正一(1982)ツヤヒサゴゴミムシダマシとナガサキトゲヒサゴゴミムシダマシの再検討. 九大農学芸誌, 36(4): 191-195.
・今坂正一・中條道崇(1983)ヒサゴゴミムシダマシ属の系統と進化(6). 月刊むし, (153): 23-26.
・Marseul, S-A. de.(1876) Coleopteres du Japon regueillis par M. Georges Lewis. Ann. Soc. Ent. Fr. (5)6: 93-142, 315-340, 447-486.
さて、ここで、ようやく、九州本土内に分布域を遮る鎖線を引いています。種の分布境界を実線で、亜種〜地域個体群の境界を鎖線で表現します。
この線のことを、分化線と呼ぶように提唱したのは、今坂(1987)が最初ですが、次に、その説明をしたいと思います。
<分化線>

分化線とは、分化型生物において、一度、ある線を越えて分布を拡げた後で、主として、地史的な要因で個体群が分断・隔離されて、いろいろな程度の種分化を起こした時の、その分布境界線を言います。
また、「分化型生物」とは、日本本土内で、想像以上に、同属の中で多くの種に異所的種分化を起こしている植物・動物の一群を言い、日浦(1978)によって提唱されました。
植物ではカンアオイ類、ササ類、テンナンショウ属など、両生類のサンショウウオ類、昆虫ではオオオサムシ属、ナガゴミムシ属、メクラチビゴミムシ類などが上げられています。
先に上げた、ヒサゴコミムシダマシ属、ジョウカイボンの大部分の属なども、同様に、加えるべきと思います。
分化線は、本来、多良山系の甲虫相の由来と起源をさぐる手法の一つとして考案しました(今坂, 1987)。
次いで、長崎県全体の甲虫相の由来を考えるために、整理・発展させました(今坂, 1989)。
その結果、種の系統や分化の程度と、分化線の位置や、複数の種による重複を解析することで、該当地域の地史を再編成できる可能性があると考えています。
・日浦 勇(1978) 蝶のきた道. 蒼樹書房,230pp.
・今坂正一(1987) 多良岳の甲虫相について. 佐賀の昆虫, (19): 261-280.
・今坂正一(1989) 長崎県の甲虫相−分化線から見た甲虫相−. 長崎県の生物: 177-184, 長崎県生物学会編.
<タラクビボソジョウカイ Hatchiana nishidai Imasaka>
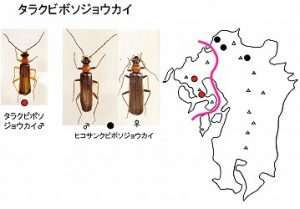
体長10mm前後、頭と上翅が黒色、前胸背と肢が赤褐色のきれいな細型のジョウカイボンで、多良山系(高来町多良岳横断林道、高来町轟の滝)と黒髪山のみで知られています。
山地帯(標高300m以上)の広葉樹林内で、林縁の葉上から見つかっていますが、非常に少なく、6♂が知られるのみで、今のところ、♀は未知です。
今坂にとって最初のクビボソジョウカイ族総説(Imasaka, 2001)において、多良山系のまとめの共著者であり、本種黒髪山産の採集者でもある西田氏に献名し、新種として記載したものです。
本種は英彦山山系に分布するヒコサンクビボソジョウカイHatchiana hikosana (Nakane et Makino)に近縁で、♂交尾器がやや異なり、肢の色が赤褐色になることで区別できます。
本種が所属するクビボソジョウカイ属 Hatchianaは、極東と北アメリカ中北部に分布域が限定され、日本に最も多くの種(12種)が分布します。Hatchiana属は3種群に分かれ、本種はその内のishiharai種群に含まれます。
ishiharai種群には5種が含まれるますが、中国地方・四国・紀伊半島に広く分布するタキモトクビボソジョウカイHatchiana takimotoana (Kiriyama)を除く4種は、ごく狭い範囲(一つの山系)に側所的に分布します。
九州の多良山系と黒髪山に本種が、
英彦山山系にヒコサンクビボソジョウカイが分布し、
四国の石鎚山系にシコククビボソジョウカイHatchiana ishiharai (M. Sato)が、
また剣山系にはサノクビボソジョウカイHatchiana sanoi Imasakaが分布します。
朝鮮半島にも、本種群に含まれると考えられるHatchianaが分布し、本州中部以東には本種群が分布しないことから、本種群もオオズナガゴミムシ種群同様、系統的には朝鮮系と考えられます。
本種はヒコサンクビボソジョウカイに最も近縁ですが、本種の方が祖先的であると考えられますので、ヒコサンクビボソジョウカイは本種から分化した種であると考えています。
結局、分化線は、多良山系と英彦山山系の間に引かれることになります。
・Imasaka, S.(2001)Taxonomic study of the genus Hatchiana in Japan (Coleoptera, Cantharidae, Podabrini). Jpn. J. syst. Ent., 7(2): 279-313.
ただ、この後も同様のことが起こりますが、多良山系と英彦山山系の間には、もう一つ、別の山系である脊振山系が存在します。
脊振山系の個体群は、種によって、多良山系と同じであったり、英彦山山系と同じであったり、あるいは、両方の種が混在していたりと、様々な性格を持っています。
最も困難なことは、脊振山系では未だ分布が確認されない種群が大変多いことです。
この場合、多良-脊振間と、脊振-英彦山間のどちらに、分化線を引くべきか、判断できないことになります。
このような場合、ここでは、便宜的に、ほとんど脊振-英彦山間に分化線を引いています。多良-脊振-英彦山間の全ての地域に分布する種群においては、脊振-英彦山間の形質の差がより大きい場合が多いという理由からです。
<マルヒサゴゴミムシダマシ Misolampidius molytopsis (Marseul)>
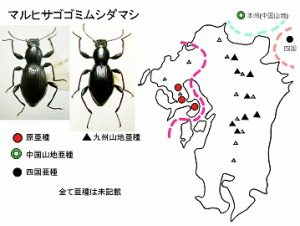
体長13mm前後、やや長めの丸っこいヒョウタン型のゴミムシダマシで、ツヤがあり後翅は退化しています。
♂前脛節中央は強く内側に張り出し、前腿節にはトゲ状の突起を持つのが特徴です。雲仙と多良山系のほぼ標高1000m以上で、キノコなどが付いた立ち枯れや倒木、樹皮下などから発見されます。ツヤヒサゴゴミムシダマシとは、標高でほぼ棲み分けていますが、標高1000m付近では同時に見られることもあります。
本種の分布域は近畿地方を除く、紀伊半島以西のブナ帯以上の山地で、♂の後脛節や交尾器の形に地域変異が見られ、紀伊半島、中国山地、四国、九州脊梁、西九州のそれぞれの個体群を区別することが出来ます(今坂・中條, 1983)。亜種の記載はまだ行われていません。
・今坂正一・中條道崇(1983)ヒサゴゴミムシダマシ属の系統と進化(5). 月刊むし, (152): 17-21.
<ウンゼンケナガクビボソムシ Neostereopalpus imasakai Nakane>
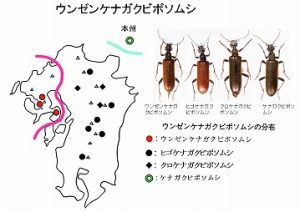
体長10mm前後、頭と前胸背、肢が黒色、上翅が褐色の細型のクビボソムシで、多良山系の佐賀県(鹿島市奥平谷)・長崎県(大村市黒木)と雲仙の山地帯(標高300m以上)に分布しています。
多良山系では渓流沿いにある広葉樹林の林縁で、シラキなどの花上、葉上で見つかりますが個体数は少ないようです(図左)。
本種は故中根猛彦博士が今坂に献名し、新種記載されたものです(中根,1983)。
雲仙ではヨドシロヘリハンミョウのところで述べた布施さん(故人)の採集品が唯一のもので、それ以外に記録されていません。
九州において、Neostereopalpus属の種としては、本種以外にも、上翅が褐色で本種に近縁と考えられるヒゴケナガクビボソムシ Neostereopalpus kyushuensis Nakane(図中左)と、
上翅が黒色のクロケナガクビボソムシ Neostereopalpus oitaensis oitaensis Nakane(図中右)、
およびその福岡亜種 Neostereopalpus oitaensis kidoi Nakaneの2種1亜種が記録されていますが、
前2種は祖母山では同一山系で見られますので、多分、別系統と思われます。
クロケナガクビボソムシはむしろ本州に分布するケナガクビボソムシ Neostereopalpus niponicus (Lewis)(図右)と同系(黒色種群)と考えています。
褐色種群では、多良山系と雲仙に分布する本種と、英彦山から九州脊梁に分布するヒゴケナガクビボソムシが、主として♂交尾器の形などで区別されますが、中間の脊振山系では本属の種は記録されていません。
・中根猛彦(1983)本邦産のクビボソムシ類について(アリモドキ科). 北九州の昆蟲, 30(1): 1-6.
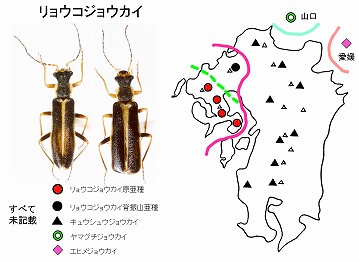
<リョウコジョウカイ Asiopodabrus sp. 8>
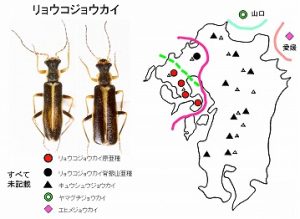
体長8mm前後、黒褐色で前胸背の周囲と上翅の合わせ目が黄褐色の、中型のニンフジョウカイです。
長崎県北部の国見山系から雲仙周辺まで、多良山系では佐賀県(鹿島市奥平谷)・長崎県(大村市南河内林道、諫早市五家原岳、高来町金泉寺、高来町轟の滝)などで記録があります。脊振山系には別亜種と思われる個体群がいます(今坂, 2002a)。
標高300m以上〜山頂付近などで見られ、春期、主としてカエデなどの落葉広葉樹の花と新芽に集まり、アブラムシなどを捕食します。
本種をはじめとするリョウコジョウカイ種群は、♂の中肢の爪が♀同様に基部に三角の付属歯を備えることにより、ニンフジョウカイ属の全ての種群から区別されます。
本種群は13種が区別可能で(2種以外は全て未記載)、福島県以西の西日本に広く分布しますが、各地に1種のみが分布し、それぞれの種は側所的に分布しており、典型的な分化型生物と言う事が出来ます。
九州産は、背振山系以西の西九州に分布する本種と、英彦山〜九州脊梁〜紫尾山まで見られるキュウシュウジョウカイに♂交尾器の形態で区別可能で、隣接する山口県にはヤマグチジョウカイ、四国の愛媛県にはエヒメジョウカイが分布します。
西九州産は、さらに、脊振山系とそれ以外で区別が可能ですが、脊振産はより、東九州のキュウシュウジョウカイに似ています。
ということで、分化線は、脊振-英彦山間に実線が、多良-脊振間に鎖線が引けることになります。
・今坂正一(2002a) 多良山系の固有種はどこからきたか. 佐賀の昆虫, (36): 481-526.
<イマサカナガゴミムシ Pterostichus imasakai Kasahara et Ohtani>
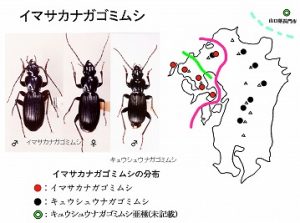
体長17mm前後、後翅が退化した黒色のゴミムシです。肢や上翅は褐色を帯びる場合が多く、広葉樹林内の渓流沿い、ガレ場などの湿った石下に生息し、比較的個体数は多いようです。
脊振山系、黒髪山(未発表)、平戸島、国見山系(未発表)、多良山系と雲仙の低地〜山頂付近に分布し、多良山系では佐賀県(鹿島市経ヶ岳、太良町中山、太良町多良岳)・長崎県(大村市黒木、諫早市五家原岳、高来町金泉寺)などで記録されています。
本種は、Kasahara・Ohtani(1988)により今坂に献名され、和名にも採用されています。
本種は英彦山〜九州山地、山口県に分布するキュウシュウナガゴミムシ Pterostichus kyushuensis Habuに近縁で、やや大型であること、肢が赤みを帯びること、♂交尾器の形などで区別されています。
記載以降、本種はキュウシュウナガゴミムシの代置種と考えていましたが、廣川(1994)により、脊振山系では混棲することが報告されています。
この結果、本来の分化線は脊振-英彦山間にあったと考えられますが、サブ的に多良-脊振間にも引いています。
・廣川典範(1994)九州における山地性ナガゴミムシについて. 月刊むし, (281): 29-31.
・Kasahara, S. & N. Ohtani (1988) Two new Pterostichus (Col., Carabidae) from the Island of Kyushu, Southwest Japan. Kontyu, 56(1): 52-61.
<タラヌレチゴミムシApatrobus kurosawai Morita>
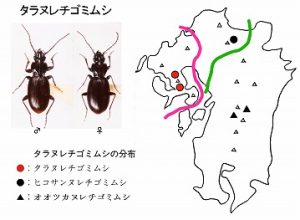
体長10mm前後、後翅が退化しややヒョウタン型をした黒色のゴミムシで、多良山系と黒髪山(未発表)の固有種です。
多良山系では、佐賀県(太良町中山)・長崎県(諫早市五家原岳、高来町金泉寺)などの山地帯(標高300m以上)に分布し、広葉樹林内の渓流沿い、ガレ場などの湿った石下に生息しています。
北九州の英彦山山系に生息するヒコサンヌレチゴミムシApatrobus hikosanus Habuに似ていますが、♂交尾器などで区別され、中間の背振山系ではどちらの種も記録されていません。
また、その後、九州脊梁の国見岳周辺でも近縁のオオツカヌレチゴミムシApatrobus ohtsukai Moritaが記載されています。
・Morita, S., 1986. A new Apatrobus (Col., Carabidae) from Mt. Tara-dake in Kyushu, West Japan. Ent. Pap. Pres. Kurosawa, Tokyo: 143-146.
・Morita, S., 1993. A new Patrobine beetle from Kyushu, West Japan. Elytra, Japan, 21(1): 173-177.
<タラダケナガゴミムシ Pterostichus taradakensis Kasahara et Ohtani>
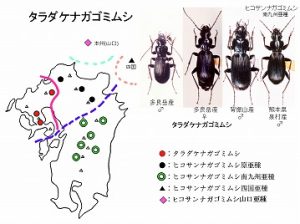
体長20mm前後、後翅が退化した黒色のゴミムシで、先のイマサカナガゴミムシよりやや大型。
脊振山系と多良山系の山頂付近に分布し、広葉樹林内の渓流沿い、ガレ場などの湿った石下に生息するが個体数は少ないようです。
多良山系では、佐賀県(太良町中山、太良町多良岳)・長崎県(諫早市五家原岳、高来町金泉寺)の記録があります。
本種は、英彦山から白髪岳までの東九州と、山口県、四国に分布するヒコサンナガゴミムシ Pterostichus hikosanus Kasaharaに近縁と考えられていますが、体型や♂交尾器で区別できます。
ヒコサンナガゴミムシはさらに、阿蘇山以南の亜種 ssp. higonis Kasahara、
四国亜種 ssp. kurosonis Kasahara、
山口亜種 ssp. kaorui Moritaなどに区別されます。
本種群の登場で、九重山系と阿蘇・祖母山系の間にも、分化線が存在することが明らかになりました。先の、ヒコサンヌレチゴミムシとオオツカヌレチゴミムシの境も、同様と考えて良いと思われます。
・Kasahara, S. & N. Ohtani (1988) Two new Pterostichus (Col., Carabidae) from the Island of Kyushu, Southwest Japan. Kontyu, 56(1): 52-61.
<タラオオズナガゴミムシ Pterostichus sp. 1>
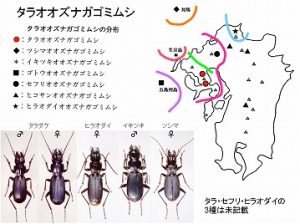
体長20mm前後、頭部と大腮が大きく、♀では左側が肥大化する。後翅が退化した黒色のゴミムシです。
佐賀県(太良町中山)・長崎県(諫早市五家原岳、高来町金泉寺)など、多良山系の山地帯(標高400m以上)に分布し、広葉樹林内の渓流沿い、ガレ場などの湿った石下に生息しますが、個体数は少ないようです。
本種を含むオオズナガゴミムシ種群は、朝鮮半島、対馬、四国、五島福江島、九州西北海上の生月島、それに九州本土各地に分布していまして、本州以北には分布しない大陸系の種群と考えられます。
本種群は上翅が短く側縁が丸い離島群と、上翅が長く側縁が直線的な九州本土群に二分され、前者は既にそれぞれ別種として区別されています。
対馬産(図右)はツシマオオズナガゴミムシPterostichus opacipennis Jedolicka、
四国産はシコクオオズナガゴミムシ Pterostichus yoshidai Kasahara、
五島福江島産はゴトウオオズナガゴミムシ Pterostichus amanoi Nakane、
生月島産(図中右)はイキツキオオズナガゴミムシ Pterostichus sakagamii Moritaです。
九州本土群は、従来、英彦山から記載されたヒコサンオオズナガゴミムシ Pterostichus macrocephalus Habu唯1種と考えられていましたが、廣川(1994, 1998)は、西九州各地の個体群は、それぞれ体型・♂交尾器など形態が異なることを指摘しました。
今坂も彼の意見を支持していて、彼の言う個体群は、山系ごとに別の種と考えています。
九州本土では、英彦山から白髪岳までの九州脊梁にヒコサンオオズナガゴミムシが分布し、
脊振山系にはセフリオオズナガゴミムシ Pterostichus sp. 2が、
多良山系(図左, 中左)に本種が分布し、
この他、北九州市産(図中央)も別の種(仮称ヒラオダイオオズナガゴミムシ Pterostichus sp. 3)と考えています。
従来、基産地の英彦山のヒコサンオオズナガゴミムシは♂が未発見で、そのために、他の産地の個体群の種名決定ができないままになっていましたが、聞くところによると、どうも、得られたらしいと言うことで、今後、整理されるだろうと思われます。
・Habu, A. (1955) Uber die Pterostichus-Arten von Berg Hiko (Die Carabidenfauna von Berg Hiko, ?). Bull. Nat. Inst. Agr. Ser. C. No. 5,: 143-156.
・廣川典範(1994)ヒコサンオオズナガゴミムシについて. 月刊むし, (285): 18-23.
・廣川典範(1998)ヒコサンオオズナガゴミムシについて(2). 月刊むし, (326): 12-14.
<ヒミコヒメハナカミキリ西九州亜種 Pidonia neglecta hizena Imasaka et Amano>
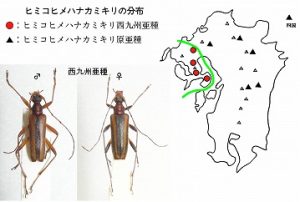
体長10mm前後、前胸背が赤褐色になる中型のヒメハナカミキリで、脊振山系の西端に位置する八幡岳と、それより西に位置するの多良山系、および雲仙岳に分布し、♂交尾器の先端にカギ状の突起があることで原亜種から区別されます。
多良山系では、佐賀県(鹿島市経ヶ岳、鹿島市奥平谷、鹿島市平谷、太良町多良岳)・長崎県(大村市南河内林道、高来町金泉寺、高来町轟の滝)など、標高350m〜1000m付近で広く見つかっています。4月中旬から6月中旬まで、カエデ、サワフタギ、ミズキなどの花上や、林縁の葉上で採集されますが、個体数は多くないようです。
西九州の個体群が、他と違うことを最初に指摘したのは天野(1996)で、雲仙産の♂交尾器先端に小突起があることに注目しています。
これを受けて、雲仙・多良両地方の個体群を精査し、今坂と共著で、亜種を記載したわけです(Imasaka & Amano, 2001)。
なお、原記載では脊振山系の東南端に位置する九千部山産1♀もパラタイプとして含めていましたが、後に入手した当地♂を精査したところ原亜種の方で、今坂(2002)により訂正しました。
結局、原亜種は背振山系の東南端に位置する九千部山と、それより東側の九州本土全域、及び四国の山地に広く分布することになり、西九州亜種は八幡岳以西の分布で、脊振山系の中に分化線が走ることになります。
主峰の脊振山は標高1055mあり、ブナも多く見られますが、今までの所、どちらも得られず、空白地帯となっています。
・天野昌次(1996) 雲仙仁田峠で採集されたヒミコヒメハナカミキリ 及び近縁の2種について. 新筑紫の昆虫, (5): 13-14.
・今坂正一(2002) 佐賀県九千部山のヒミコヒメハナカミキリ. 月刊むし, (376): 52-53.
・Imasaka, S. & M. Amano (2001) Pidonia neglecta hizena n. subsp., a new subspecies from Western Kyushu, Japan (Coleoptera, Cerambycidae). SUKUNAHIKONA, Spec. Publ. Japan Coleopt. Soc. Osaka, (1): 337- 342.

