<島原半島>
雲仙噴火で知られる島原半島は、長崎県南部の有明海に突き出る島嶼性の強い半島で、長崎市と熊本市のほぼ中間に位置しています。
東西25km、南北32km、面積463k?で、おおまかに言うと東京23区や、屋久島、西表島などとほぼ同じ広さがあります。
地形は起伏に富み、中央部に雲仙火山の普賢岳(1,360m)を始めとして多くの山が連なり、平地に乏しい地形です。
1990年以降の噴火活動により、普賢岳東斜面より出現した火山ドームはその後成長を続け、標高1,470mの平成新山を形成したことは記憶に新しいと思います。
(島原半島の固有種)
1975年冬に帰郷して以来、家業の呉服業を手伝う傍ら、昼休みや終業後、休みなどに寸時を惜しんでカミキリとハムシの採集を行ないました。
しかし、日帰りで行ける範囲はごく狭く、瞬く間に限界を感じ、採集対象を甲虫類全体に広げました。
採集した甲虫は標本を作り、可能な限り同定して毎日記帳しました。
当時の記録を見ますと、1年目には海外遠征などを除いてのべ115回、2年目も133回、3年目でも88回と、シーズン中はほぼ毎日、30分でも、屋外で虫を採っていたことが伺えます。
そのうち標本箱とノートが溜まり、1977年からは、採集結果を地元の同好会誌などに投稿しました。
専門家から同定誤りを指摘され、同定方法を教わって訂正し、特殊な採集法も教えてもらって実行しました。
そして、そんなことを繰り返し、地元の甲虫なら、何が、どこに、いつ頃いるかを言えるようになったのは、10年ほど経った頃でしょうか。
<島原半島の甲虫>
ほぼ20年間調査を続けて、1995年に久留米へ転出したのを機会に総まとめをし、「島原半島の甲虫相」1〜5(今坂 1999〜2002b)として、島原半島産甲虫102科1,972種を整理・記録しました。
ここに図示しているのは、島原半島の中核にあたる雲仙岳の標高1000m以上に生息している甲虫67科674種のうち、特徴的な種を並べたものです。
(雲仙岳の標高1000m以上に生息している甲虫)

雲仙岳の甲虫相については、今坂(2006)に詳しく、別に以下のホームページに種リストと共に掲載していますので、詳細はそちらをご覧下さい。

島原半島では本州、四国、九州に生息するいわゆる本土系の種をベースとして、オオセンチコガネ、ドウイロチビタマムシなどの大陸や北方からの広域分布種と、サンカクスジコガネ、タテスジフトカミキリモドキなど、琉球や南方からの広域分布種が多いという特徴がみられました。
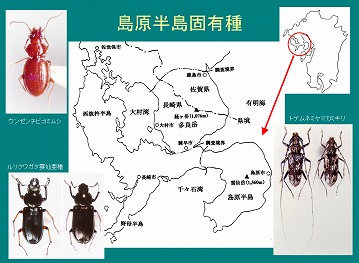
固有種として、ウンゼンチビゴミムシなど3種と、ルリクワガタ亜種(ウンゼンルリクワガタ)が発見されました。
ファウナ調査が進んでいる神奈川県や、国内産リストなどを参考にしますと、最終的には500k?足らずのこの半島に、3,000種程度の甲虫が生息していると予想されますので、ようやく、2/3程度を実際に記録することが出来たことになります。
<多良山系の甲虫>
多良岳は島原半島の北部に隣接し、長崎と佐賀の県境を跨いでそびえる山系です。主峰の経ヶ岳(1,076m)を中心としたほぼ500k?の楕円形で、山系の範囲は島原半島とほぼ同じ程度の広さがあります。
当然、採集した大部分の種は島原半島と共通ですが、ほんの隣の陸続きの山系であるにもかかわらず、クロツヤハネカクシ、マダラクワガタ、ムラサキツヤハナムグリ、ベニヒラタムシなど、島原半島ではいくら探しても採集できないような種も見つかります。
(多良山系固有種)
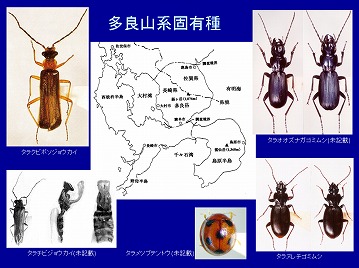
1987年に、長崎県側のほぼ10年分の成果と、緒方・西田両氏による佐賀県側の調査結果を合わせて、多良山系のまとめを行い、93科1,469種を記録しました(今坂・緒方・西田 1987)。
さらに2002年には、その後15年間の採集記録と文献記録を整理して438種を加え、改めて多良山系産甲虫として101科1,901種を報告しました(今坂・西田 2002)。
多良山系の甲虫相の特徴は、島原半島同様、本州、四国、九州に生息する本州系の種をベースとしますが、さらに北海道まで分布する北方系の種が多く、南方系の種は少ないのが特徴です。
タラヌレチゴミムシ、タラクビボソジョウカイなどの多良固有種も数種発見されています。
<島原半島と多良山系の分布型によるファゥナの比較>
島原半島と多良山系はほぼ同じ地理的位置にあり、面積、標高、地形や、植生などの環境もほとんど変わりません。
記録された種もほぼ同数で、その2/3が互いに共通種です。
ということは、逆に、それぞれ1/3は別の種が得られていることになりますが、単なる偶然や、採集者が違うこと、あるいは採集方法や時期が違うことなど、調査精度の違いが原因の大半と思われます。
それらの誤差を補正して考えると、実際は85%程度が共通になり、それでも、それぞれ15%程度は違った種が分布することになると考えられます。
両地域のファウナの比較をする場合、種の直接比較では、それ以上の考察は得られません。
ここはまた、別のファクターを導入して、別の側面から比較する必要があります。
結局、種が抱えている要素のうち分布パターンに注目し、島原半島と多良山系に分布するそれぞれの種を、便宜的に、次15のタイプに区分してみました。
A: 北方系広域分布種・・・・・・・北海道、本州、四国、九州、(屋久島、トカラ列島)、朝鮮半島、(中国大陸、台湾)
B: 南方系広域分布種・汎世界種・・北海道、本州、四国、九州、琉球列島、(朝鮮半島、中国大陸、台湾)、汎世界
C: 大陸・本土系種・・・・・・・・本州、四国、九州、(屋久島、トカラ列島)、(朝鮮半島)、中国大陸、(台湾)
D: 大陸系種・・・・・・・・・・・九州、(屋久島)、朝鮮半島、中国大陸、(台湾)
E: 北方・本土系種・・・・・・・・北海道、本州、四国、九州、(屋久島、トカラ列島)
F: 本土系種・・・・・・・・・・・本州、四国、九州、(屋久島、トカラ列島)
G: 暖地系種・・・・・・・・・・・本州、四国、九州(以上の山地を除く)、琉球列島、(台湾)
H: 琉球系種・・・・・・・・・・・(四国)、九州(山地を除く)、琉球列島、(台湾)
I: 九州・四国固有種・・・・・・・四国、九州、(屋久島)
J: 九州固有種・・・・・・・・・・九州、(屋久島)
K: 長崎・佐賀固有種・・・・・・・長崎県、佐賀県
M: 分布不明種・・・・・・・・・・未記載種など、分布域不明
O: 北九州固有種・・・・・・・・・九州北半分
P: 背振・多良山系固有種・・・・・多良山系・背振山系
Q: 多良山系固有種・・・・・・・・多良山系
そして、それぞれの種数の全体に対する比率を算出してみました。
( )内は、分布していない場合もあることを示しています。
<多良山系における分布型構成の年次比較>
多良山系では、1987年に行った第一次のまとめの時点(今坂・緒方・西田,1987: 1469種)では、
F: 本土系の比率が最も高く、
次いで、E: 北方・本土系、A: 北方系広域分布、B: 南方系広域分布、C: 大陸・本土系の順に減少していき、
より南方系のG: 暖地系やH: 琉球系、固有種のJ: 九州固有、I: 九州・四国固有、D: 大陸系、K: 長崎・佐賀固有など特殊な分布を示す種はかなり低くなつています。
(表-1)
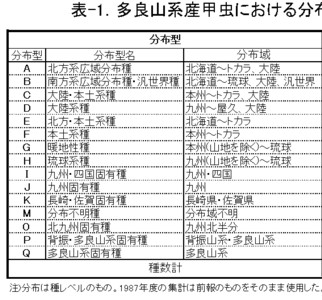
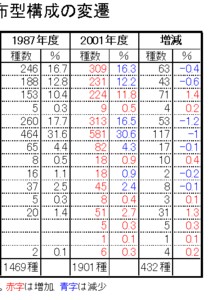
これらの比率は、438種を追加した2002年のまとめの際(今坂・西田,2002: 1901種)にも再び算出しましたが、
新たに設定したO: 北九州固有、P: 脊振・多良固有、Q: 多良山系固有が加わっただけで、順位は変わらず、
また、誤差も最大で1.5%程度と、1987年とほとんど有意の差はみられませんでした。
これらのことから、調査精度にかかわらず、同じファウナを持つ地域の分布型の比率はほぼ一定で、表-1に示した比率は、多良山系の甲虫相の特徴を表していると判断されます。
<多良山系と島原半島の分布型比較>
同様に、多良山系(2001年)と島原半島を比較してみます。
(表-2)
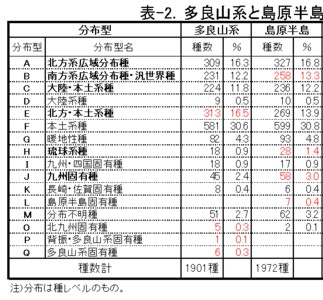
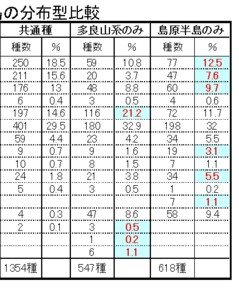
多良山系と島原半島の共通種1353種においては、F: 本土系種が最も多く、次いでA: 北方系広域分布種、B: 南方系広域分布種、E: 北方・本土系種、C: 大陸・本土系種などが多く、以上の合計が91.2%を占めています。
次いで、G: 暖地系種、J: 九州固有種、I: 九州・四国固有種、H: 琉球系種など。僅かにD: 大陸系種、K: 長崎・佐賀固有種、O: 北九州固有種や、一部にM: 分布不明種も含まれていました。
以上の種は、多良山系、島原半島の双方に分布しており、両地域の甲虫相の基盤になる種群であり、比較的広域分布種の割合が多いようです。
一方、多良山系のみで記録された種は547種であり、隣の島原半島のみで記録された種は618種です。
表-2のうち、お互いの分布型に含まれる種の比率を比較して、1%以上多いか、片方だけに見られる分布型の%のバックの色を薄青色にしてみました。
この結果、多良山系ではE: 北方・本土系種が、島原半島と比較して特に多いことが解ります。
例えば、アイヌハンミョウ、ガロアミズギワゴミムシ、サドモリヒラタゴミムシ、サワダマメゲンゴロウ、マダラクワガタ、ムラサキツヤハナムグリ、イガラシカッコウ、ベニヒラタムシ、ヨツボシオオキノコムシ、ムネアカナガクチキムシ、オオモンキゴミムシダマシ、セスジヒメハナカミキリ、ホソトラカミキリ、クロニセリンゴカミキリ、マルモンタマゾウムシなどが相当し、これらは、島原半島では記録されていません。
そして、多分、その多くが実際に、分布していないだろうと推定されます。
その他の分布型では、島原半島のみとほぼ同じか、逆に少ないという結果になっています。
島原半島には分布しない、Q: 多良山系固有種であるタラヌレチゴミムシ、タラオオズナガゴミムシ、タラダケオノヒゲアリヅカムシ、タラダケヒゲナガアリヅカムシ、タラチビジョウカイ、タラクビボソジョウカイ、
O: 北九州固有種であるサイカイヒサゴアリヅカムシ、ヒゲブトムネトゲアリヅカムシの一種?、オオトラフコガネの一種、
P: 背振・多良山系固有種であるタラダケナガゴミムシ、
K: 長崎・佐賀固有種であるヒゲブトムネトゲアリヅカムシの一種?、マツウラヒゲナガアリヅカムシ、ナガサキトゲヒサゴゴミムシダマシなどが分布することが特徴です。
一方、島原半島では
A: 北方系広域分布種であるカワラハンミョウ、オオセンチコガネや、
C: 大陸・本土系種であるルイスハンミョウ、ドウイロチビタマムシ、
B: 南方系広域分布種であるセマルオオマグソコガネ、ヘリグロテントウノミハムシなどや、
H: 琉球系種であるサンカクスジコガネ、タテスジフトカミキリモドキなど、
それにJ: 九州固有種のウンゼンクビボソジョウカイなど、種々の分布型について、それぞれ0.9〜3.9%の範囲で多良山系より比率が高いという結果になっています。
このうち、最も多いのはB: 南方系広域分布種で、多良山系のみより27種(3.9%)も多くなっています。
また、当然ながらL: 島原半島固有種も7種(1.1%)分布しています。
多良山系では、飛び抜けて北方・本土系種が多くなり、それに対して、島原半島ではさまざまなルートで侵入した広域分布種が多く、中でも暖流の影響を受けた南方系種の割合が高いと言えると思います。
<多良山系と島原半島の甲虫相の違い>
つまり、多良山系と島原半島の甲虫相の相違は、九州脊梁から英彦山経由で分布を拡げてきた山地性の種が、どの程度侵入できたかどうかに掛かっているようです。
多良山系では比較的多くの種が、陸づたいに分布を広げ、その甲虫相を構成したのに対して、島原半島ではかなりの割合の種が到達することが出来ず、甲虫相は全体として調和を欠いた状態に陥ったものと考えられます。
そのために空白となった生態的な隙間には、主として移動力の大きい南方系・北方系両方の広域分布種が、海流分布やその他の手段によって、多く侵入したのでしょう。
南方系広域分布種の一部は、多良山系へも侵入したと思われますが、島原半島と比較して、暖流から隔てられた多良山系には、多くは到達・定着できなかったものと考えられます。
引用文献
今坂正一(1987) 多良岳の甲虫相について. 佐賀の昆虫, (19): 261-282.
今坂正一(1999) 島原半島の甲虫相1. 長崎県生物学会誌 (50): 125-170.
今坂正一(2000) 島原半島の甲虫相2. 長崎県生物学会誌 (51): 19-39.
今坂正一(2001a) 島原半島の甲虫相3. 長崎県生物学会誌 (52): 56-73.
今坂正一(2001b) 島原半島の甲虫相4. 長崎県生物学会誌 (53): 65-84.
今坂正一(2002a) 多良山系の固有種はどこからきたか. 佐賀の昆虫, (36): 481-526.
今坂正一(2002b) 島原半島の甲虫相5. 長崎県生物学会誌 (55): 53-73.
今坂正一(2006) 雲仙岳の甲虫相. 雲仙岳の生物: 101-115, 長崎新聞社刊, 長崎県生物学会編.
今坂正一・西田光康(2002) 多良岳の甲虫相2001. 佐賀の昆虫, (36): 389-480.
今坂正一・緒方 健・西田光康(1987) 多良岳の甲虫相. 佐賀の昆虫, (19): 176-260.