
もう10年ほど前にやった調査ですが、何時までも記録しないままでは良くないので、思い出しながら書いています。
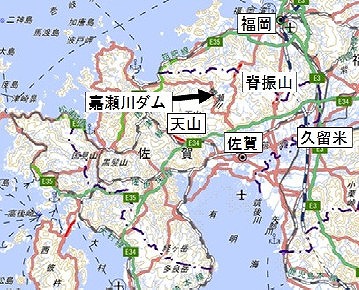
嘉瀬川ダム(標高304m)は、脊振山系の山間、佐賀県のほぼ中央部である佐賀市の、ほぼ20km北方にあります。
このダムは、佐賀市を縦断する嘉瀬川の上流に位置し、下流に広がる佐賀平野の洪水被害の軽減、農業用水・飲み水など水道水の確保を目的として建設されました。
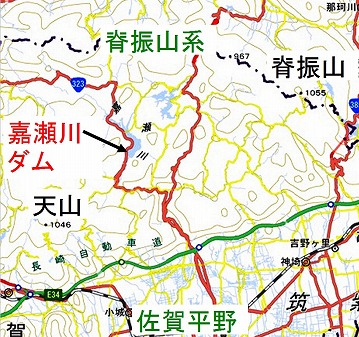
1973年より計画され、2005年よりダム本体の建設を開始し、2010年10月には試験湛水を開始、2012年2月に湛水を完了し、同年3月に竣工したものです。
この最終段階の2009年5月から2012年11月までの4年間、ほぼ春、夏、秋の3季、のべ11回(2010年夏は中止)、ダム内とその周辺で昆虫類の調査を行いました。
ここでは、そのうち、私が担当した甲虫類について報告したいと思います。
調査に参加したのは、計画された祝さんを筆頭に、古川、田畑、大城戸、平原、今坂の6名です。
採集データとしては、採集者を区別していませんので、この全員で採集したことになります。
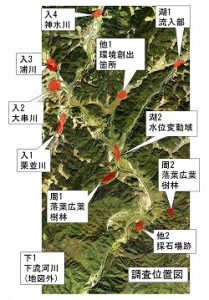
調査地点は以下のとおりです。
嘉嘉湖P1: 嘉瀬川ダム湖への本川流入部。本川の上流部という扱い。
嘉嘉湖P2: 水位変動域。河川からダム湖に変化していく場所の代表地点。川岸、湖岸に調査地点を設置するため水位が上昇するにつれて地点を右岸側の道路に沿って変更しました。
嘉嘉入P1: 栗並川。嘉瀬川の流入河川。
嘉嘉入P2: 大串川。嘉瀬川の流入河川。
嘉嘉入P3: 浦川。嘉瀬川の流入河川。
嘉嘉入P4: 神水川。嘉瀬川の流入河川。
嘉嘉周P1: 嘉瀬川の周辺環境として設置。ダム湖試験湛水の影響は直接は受けない場所。場所は右岸の境谷。環境は細流沿いに植林、落葉広葉樹、常緑樹があります。
嘉嘉周P2: 嘉瀬川の周辺環境として設置。ダム湖試験湛水の影響は直接は受けない場所。場所は左岸の西畑瀬。環境は常緑樹、落葉広葉樹。
嘉嘉他P1: 環境創出箇所の音無地区。河川水辺の国勢調査でのその他の環境として地点を設定しました。
嘉嘉他P2: 採石場跡。採石場跡が水位変動域となっていたため、そこで調査を行いました。河川水辺の国勢調査でのその他の環境として地点を設定してあります。
嘉嘉下P1: 嘉瀬川の本川の下流部。ダム直下は工事で立ち入りできなかったので、少し下流側にずらして古湯の貝野川合流部で調査を行いました。
(訂正.
当初、川の右岸と左岸を、川の下流から上流を眺めて書いておりました。正しくは、川の上流から下流を眺めて、右側を右岸、左側を左岸と呼ぶそうです。全て訂正しました。ご教示いただいた山元さんにお礼申し上げます。)
上記の地点表記は、国土交通省の河川水辺の国勢調査のダム版の表記に合わせていますが、ここでは煩雑なために、この後は嘉嘉湖P1を嘉嘉とPを略して、湖1、入1のように表記します。
調査の大部分は任意採集の見つけ採りですが、2009年度と2011年度はライトトラップ、ベイトトラップも行いました。
調査地点ごとに、確認された種とその推移について見ていきたいと思います。
1. 湖1(佐賀市富士町大字関屋、標高306m)
嘉瀬川本流のダムより上流部、ダム本体より4kmほど上流の地点です。左岸の高水敷は公園化され、サクラやカエデなどが植栽されています。河川敷は礫混じりの砂地で、岩もゴロゴロしていました。右岸側は藤瀬神社の広葉樹の社寺林が隣接し、その北側は集落と耕作地が広がっています。

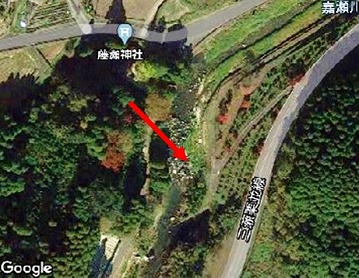
建設以前の航空写真は元図の縮尺がでかいので、ごく粗い写真になります。現在の航空写真はグーグルから引用しました。以下も同様です。当地点は、調査前と終了時は、ほとんど様子は変わっていません。
調査の様子は以下の写真の通りです。



湖1では任意採集として、2009年5月5, 9日、8月18日、2010年5月1日、10月1日、2011年5月7日、8月29日、11月7日、2012年5月24日、8月25日、10月31日の10回、ライトトラップとピットフォールトラップを2009年5月2-3日と2011年5月4-5日の2回実施しています。
湖1では4年間の調査で177種が確認できました。
年度毎では、2009年(60種)、2010年(54種)、2011年(71種)、2012年(92種)です。
このうち、以下の9種は佐賀県から記録の無い種で、佐賀県初記録になります。
なお、佐賀県産、および、脊振山系産甲虫の記録の有無については、西田光康氏(武雄市)にご教示いただきました。厚くお礼申し上げます。
ヒメシジミガムシ、ユミセミゾハネカクシ、アシマダラカワベメダカハネカクシ、ヨコモントガリハネカクシ、ウエダニンフジョウカイ、トゲアシチビケシキスイ、クサイチゴトビハムシ、イヌノフグリトビハムシ、タデトゲサルゾウムシ
(ウエダニンフジョウカイ♂背面、♂交尾器腹面)

本種は本州(山口),九州(熊本県北部以北)に分布し、個体群によって、白っぽかったり、黒ずんだりします。久留米の低山地では本種が圧倒的優占種です。
一方、長崎県本土全域と佐賀県西部には、未記載種のナガサキニンフジョウカイが分布し、本種と棲み分けています。
(ナガサキニンフジョウカイ♂背面、♂交尾器腹面)

嘉瀬川ダム産も佐賀県産の多くと同様に、当初は、長崎産の感覚で、ナガサキニンフジョウカイと誤同定していました。それもあって、佐賀県ではウエダニンフジョウカイは未記録になっています。
なお、ウエダの♂交尾器側片の腹側突起(矢印)は太く、肩が丸く張って直線的ですが、ナガサキでは細く鋭角的で波打った形です。
(モロツカニンフジョウカイ♂背面、♂交尾器腹面)

さらに、宮崎・熊本両県の九州脊梁沿いにはモロツカニンフジョウカイがいて、この種はナガサキに良く似ていますが、♂交尾器側片の腹側突起(矢印)はより太く、側方の抉れもやや深いです。
(ウエダ、ナガサキ、モロツカの3種のニンフジョウカイの分布概要)
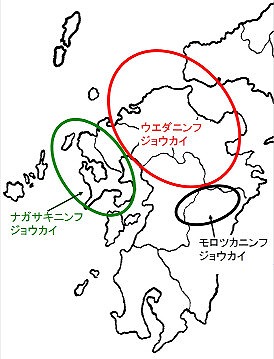
佐賀県では、ヒミコヒメハナカミキリや、リョウコニンフジョウカイの例にもあるように、脊振山系以東と、八幡岳以西で、亜種や種を分ける例が数例あり、唐津から多久に抜ける線は、佐賀県の生物地理上、最も重要な分化線になりそうです。

本種はオドリコソウチビケシキスイと共にオドリコソウの花にいます。トゲアシの方が、点刻がかなり小さく、ツヤがあるように見えます。本種の方が、山地ではやや優勢で、オドリコソウの方はむしろ低地に多いようです。


2種共にタデ科に多く、特に湿地のミゾソバに多く見られます。前者は図鑑に図示されていないので、記録が多くありません。

草地で見つかりますが、ホストのイヌノフグリ葉上では見たことがありません。
また、以下の21種は、脊振山系から記録の無かった種で、脊振山系初記録となります。
ニセヒメユミセミゾハネカクシ、ナミフタホシメダカハネカクシ、キアシチビコガシラハネカクシ、ナラノチャイロコガネ、タテスジナガドロムシ、タマムシ、キイロニンフジョウカイ、モリモトクロチビジョウカイ、クロアオケシジョウカイモドキ、アカボシチビヒメハナムシ、キムネヒメコメツキモドキ、クロヒメハナノミ、アズキマメゾウムシ、ヒメコブハムシ、ムネアカキバネサルハムシ、ツヤキバネサルハムシ、アカバナトビハムシ、クビボソトビハムシ、ニセチビヒョウタンゾウムシ、ダイコンサルゾウムシ、アオバネサルゾウムシ。

春に河川敷などで見られますが、余り多くありません。
(モリモトクロチビジョウカイ♂背面・側面)

かつて、(仮称)シマバラチビジョウカイとして記録していた種ですが、森本先生の逝去後、同じ弟子仲間である高橋直樹博士と共著で、新種記載をするのに当たって、種小名にmorimotoiと献名し、和名も変更しました(Takahashi, N. & Imasaka, 2020)。
(モリモトクロチビジョウカイ♂腹面と♂腹板拡大図)

♂腹節腹板の末端節はY字状に広がっています。近似のヒコサンクロチビジョウカイに似ていますが、Y字部分はより細く、華奢な感じです。
最初に島原半島で見つけたので、こう仮称していました。本種は九州全域と、下関で記録されています。
高橋さんが、Malthodesの複雑な♂腹節背板の構造に対して、対応する構造が♀の腹節背板に存在することを、初めて発見して、詳しく解説した報告になっています。
Takahashi Naoki & Shoichi Imasaka, 2020. Two new species and a new distribution record of the genus Malthodes Kiesenwetter (Coleoptera: Cantharidae) from Japan, with notes on the male and female abdominal morphology. Zootaxa, 4778(2): 357-371.

久留米あたりに多いケシジョウカイモドキは、前胸は点刻とシワでザラザラしていて光沢はありませんが、本種は点刻は少なく、ツヤがあります。時々採れますが、かなり少ないです。
(前胸背、左: クロアオケシジョウカイモドキ、右: ケシジョウカイモドキ)

(ニセチビヒョウタンゾウムシ、赤矢印は外側に丸く張り出した口吻先端部)

近似のチビヒョウタンゾウムシは、樹林林床の落ち葉篩いで比較的普通に採れます。
本種はめったに得られず、かなりの珍品と思っていましたが、当調査では、川沿いの草地の複数地点で採れていて、そのような環境が本種の生息地である可能性を感じました。
触角付け根付近で、外側に丸く張り出すのが本種で、チビヒョウタンゾウムシは口吻付け根からほぼ直線的に同幅で、側方に張り出さないので区別できます。
その他に、この地点を特徴づける種としては、以下の種があります。

アイヌハンミョウは、河川の砂地に特有の種で、県内の産地は多くありません。脊振山系、作礼山、多良山系などで記録されている程度です。
九州でも福岡・熊本・宮崎の各県ではRDB種に挙げられています。嘉瀬川ダムでは他にも入2、入4、湖2で見つかっています。
嘉瀬川ダムを含む佐賀地方では、2010年7月24日に「平成21年7月中国・九州北部豪雨」と呼ばれる豪雨災害のため、湖1の本種の生息地を含む河川敷の砂地は大半が流失しました。その為、2011と2012年度は確認出来ませんでした。
ジュウシホシツツハムシは草原性の種ですが、比較的狭い草地でも見られることがあります。フトキノカワゴミムシ、コブノコギリゾウムシ、アカナガクチカクシゾウムシの3種と共に、当地点のみで見つかっています。
湖1で確認された177種について、生態的に依存する環境を見てみますと、
例えば、セボシジョウカイであれば、広葉樹林、草地、河川、湿地、耕作地の5区分で出現しますので、按分して各区分1/5ずつ。アカナガクチカクシゾウムシであれば、広葉樹林のみで1、フタモンクビナガゴミムシであれば、河川と湿地で、各区1/2ずつということになります。
出現種の全てにこの生態的に依存する環境の値を振り分けて、その集計をすると、地点ごとの特徴が出ます。
湖1では、広葉樹林51と1/3、針葉樹林4と1/3、草地32と8/9、裸地2と3/4、河川50と1/2、湿地19と2/9、耕作地16と集計されました。
これを総出現数177で割ると、生態要素としては、広葉樹林28.98%、針葉樹林2.45%、草地18.58%、裸地1.55%、河川28.51%、湿地10.86%、耕作地9.07%になります。
つまり、この湖1地点の特徴としては、広葉樹林と河川の要素がそれぞれ3割弱で最も大きく、次いで、草地が2割弱、湿地と耕作地が1割程度、針葉樹と裸地は2-3%でごく少ない、ということになります。
これを年度別に集計してみると、以下のようになります。
2009年度は30.89%、1.67%、18.53%、2.22%、27.69%、10.06%、8.94%
2010年度は22.69%、2.47%、20.22%、1.08%、34.72%、10.80%、8.02%
2011年度は26.38%、1.41%、20.63%、1.64%、26.15%、12.30%、11.48%
2012年度は31.68%、1.63%、20.27%、1.00%、25.89%、10.49%、9.04%
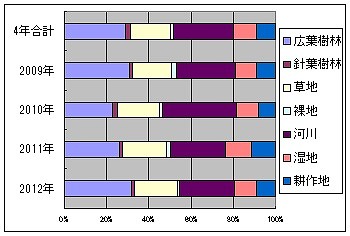
2010年度に広葉樹林の比率が多少下がり、その分、河川の比率が増えていますが、その他の年度ではほぼ変化がありません。
さらに条件を少し厳しくして、1つあるいは2つの環境のみに依存しているものだけを抽出して集計してみましょう。そうすると、限られた環境でしか出現しない種が残ります。
湖1ではこうした依存環境限定種が、出現種177種のうちのほぼ半数、87種でした。
そこから算出した比率は、45.40%、4.60%、12.07%、0.57%、29.31%、6.32%、1.72%となりました。
つまり、広葉樹林の比率がかなり上がり、河川は微増、耕作地は激減し、草地と湿地は6割程度に減少しています。
湖1の出現種全体の比率より、こちらの方が、より年度ごとの地点の特徴が出るように思われます。
それでは、依存環境限定種で同様に計算して年度ごとの比率を出してみましょう。
2009年度は56.52%、4.35%、6.52%、2.17%、26.09%、2.17%、2.17%
2010年度は32.69%、3.85%、17.31%、0.00%、42.31%、3.85%、0.00%
2011年度は43.55%、3.23%、16.13%、0.00%、25.81%、8.06%、3.23%
2012年度は51.22%、3.66%、13.41%、0.00%、24.39%、4.88%、2.44%
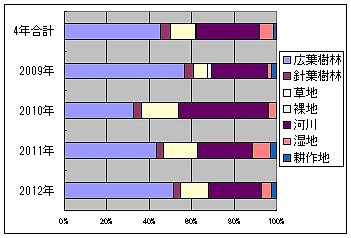
やはり、2010年度の比率は別にして、大きな変化はありません。ただ、河川と湿地要素は徐々に減っていることがうかがえます。
結論として、調査終了時も開始時と生態環境としてはそれほど変化が無いことが解ると思います。
この地点の700mくらい下流に堰があり、この地点まではダム湖の水位は上がってこないので、地点そのものの、見かけの環境は変化していません。ただ、アイヌハンミョウが2011年以降確認できなくなったことから、河川敷内については何らかの変化が起こった可能性もあります。
次に、ダム底に沈んでしまって、消失した場所について見てみましょう。
2. 湖2 (元、佐賀市富士町大字栗並、推定標高270m)
ダム湖となった富士しゃくなげ湖のほぼ中央、銀河大橋のほぼ直下にあたる地点です。調査時は河川の周囲は草地や裸地が大部分でしたが、ダム建設以前は河川沿いに集落や耕作地が広がっていたと想像されます。
ちょうど嘉瀬川上流部と、支流の神水川の合流地点に当たり、河川敷はかなり広く砂に被われていて、アイヌハンミョウを主とした、砂地の河川敷特有な種の良い生息地になっていました。



湖2では任意採集として、2009年5月5日、8月19日、2010年5月1日、9月29日、2011年5月6日、8月28日、10月23日、2012年5月24日、8月26日の9回、ライトトラップとピットフォールトラップを2009年5月2-3日と2011年5月4-5日の2回に実施しています。
湛水の影響で2011年5月までで河川敷の調査は出来なくなり、その後は、国道からその地点に降りる道沿いの採集になり、出現種構成は完全に変化しました。


湖2では4年間の調査で174種が確認できました。年度ごとでは、2009年(91種)、2010年(52種)、2011年(75種)、2012年(41種)です。
このうち、以下の12種は佐賀県から記録の無い種で、佐賀県初記録になります。
イグチマルガタゴミムシ、ユミセミゾハネカクシ、ホソフタホシメダカハネカクシ、ツユキクロホソジョウカイ、ウエダニンフジョウカイ、クサイチゴトビハムシ、アサトビハムシ、ヌカキビタマノミハムシ、カナムグラヒメゾウムシ、ミズキトゲムネサルゾウムシ、タデトゲサルゾウムシ、フタキボシゾウムシ



以上3種はそれぞれ、砂礫混じりの河川敷、渓谷沿いの樹林、河川のヤナギに見られる種で、河川環境がないと生息できない種です。
(アサトビハムシ、前頭隆起の後の溝(赤矢印)がクッキリしているのが特徴)



以上3種は、耕作放棄地など、荒れた草地などで見られます。ホストの名がそのまま種名になっています。

本種はミズキの葉上にいる未記載種です。
また、以下の19種は、脊振山系から記録が無く、脊振山系初記録になります。
ヒメツヤマルガタゴミムシ、マグソガムシ、ヤマトニセユミセミゾハネカクシ、ナミフタホシメダカハネカクシ、キアシチビコガシラハネカクシ、クロガネトガリオオズハネカクシ、ヒロエンマアリヅカムシ、コウゾチビタマムシ、キイロニンフジョウカイ、アカボシチビヒメハナムシ、ニセミツモンセマルヒラタムシ、トケジクロヒメハナノミ、アズキマメゾウムシ、キイロナガツツハムシ、ムネアカキバネサルハムシ、サシゲトビハムシ、ヨモギトビハムシ、ダイコンサルゾウムシ、マダラアシゾウムシ。
その他に、この地点を特徴づける種としては、以下の種があります。

アイヌハンミョウは、先に述べたように、嘉瀬川ダムの最も特徴的な種ですが、この地点が調べた中では最大の生息地でした。
湖1の項で、2010年7月24日の「平成21年7月中国・九州北部豪雨」で河川敷の砂地が消失したことを述べましたが、湖2ではこれより後の2011年5月4-6日が最終の確認ですから、湖2の生息地はダメージを受けたとしてもなんとか生息し続けていたものと思われます。しかし、その次の調査日8月28日以降は湛水により水没し、生息地は消滅しました。
本来なら、この生息地が水没しても、その他の中小の生息地が存続すれば継続できたと思われますが、その直前に大雨で中小の生息地が流出し、最大の生息地が水没したことで、あるいは嘉瀬川水系のアイヌハンミョウは姿を消した可能性があります。
細かい砂地に生息するため、山地では谷間の盆地的な場所が砂の堆積地になり、本種の良好な生息地になります。生息地の多くはダム湖に利用しやすい地形なので、常に消滅の危険性を秘めています。
カワチマルクビゴミムシ、オオマルクビゴミムシ、キアシマルガタゴミムシは3種共に、砂礫質の河川敷の水辺に見られ、当地点と下流の下1のみで確認されています。特にオオマルクビゴミムシは佐賀県内では脊振山系のみで記録されているので重要です。

また、以下の13種は湖2のみで確認されました。既に脊振山系からは記録されています。
ヒョウゴミズギワゴミムシ、カワチゴミムシ、マルガタツヤヒラタゴミムシ、クビボソゴミムシ、オオナガヒラタコメツキ、クリイロアシブトコメツキ、ミズギワコメツキ、キイロセマルキスイ、マルクビツチハンミョウ、ハイイロヤハズカミキリ、フタイロセマルトビハムシ、チャバネツヤハムシ、ハイイロトゲトゲゾウムシ


この2種も砂礫質の河川敷に見られ、前者は水辺に、後者は草付の石下などに見られます。
湖2で確認された174種について、生態的に依存する環境を見てみますと、湖2では、広葉樹林50と2/3、針葉樹林1と2/3、草地31、裸地2と4/7、河川51と1/4、湿地17と3/4、耕作地19と集計されました。
これを総出現数174で割ると、生態要素としては、広葉樹林29.12%、針葉樹林0.96%、草地17.82%、裸地1.48%、河川29.45%、湿地10.20%、耕作地10.97%になります。
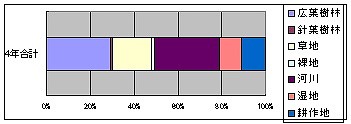
先の湖1と、各区分とも殆ど同様の比率になったのが非常に興味深いです。
つまり、この湖2地点の特徴としては、広葉樹林と河川の要素がそれぞれ3割弱で最も大きく、次いで、草地が2割弱、湿地と耕作地が1割程度、針葉樹と裸地は1-2%でごく少ない、というところまで同じです。
嘉瀬川ダム周辺の川沿いで調査すると、ほぼ同様の比率になるのでしょうか?
他の地点でも検証してみたいと思います。
湖1の場合、出現種全体よりは、依存環境限定種の方が、より年度変化が顕著に表れることが解ったので、湖2でも、年度変化はこちらで見てみましょう。
依存環境限定種は湖2では91種で、43.96%、1.10%、13.19%、0.00%、31.87%、4.95%、4.95%です。
2009年度は25.61%、0.00%、12.20%、0.00%、48.78%、7.32%、6.10%
2010年度は51.79%、1.79%、10.71%、0.00%、32.14%、3.57%、0.00%
2011年度は31.94%、0.00%、12.50%、0.00%、41.67%、8.33%、5.56%
2012年度は60.00%、2.50%、15.00%、0.00%、12.50%、2.50%、7.50%
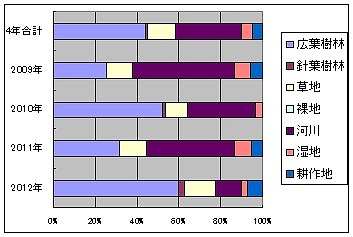
2010年度と2012年度に、広葉樹林の比率が倍増し、河川の比率が2/3に下がっていますが、1つには、この2回はライトトラップとピットフォールトラップを行わなかったためと考えられます。
これらのトラップを行った2009年度と2011年度は、比率の概要はほぼ同様であることから、2011年時点まではほぼ同様の環境が保たれていたことが推定できます。
これが2012年度になると、湛水で河川敷は消滅して、調査場所が道路沿いに変わり、確認される種数は半減しています。
特に河川要素は48.78%から12.50%へと、1/4ほどに減少著しく、水物等、湿地要素も1/3になっています。草地と耕作地は微増で、広葉樹林が倍増しているのが顕著です。この事は林縁のみの調査になったことを反映しているものと思われます。当然ながら、調査環境は激変しています。
以上、依存環境限定種による比率の比較で、調査地の年度ごとの環境変化が、ある程度掴めることが解ったので、その他の地点でも計算してみたいと思います。
つづく

