

2008年12月25日に、陸生ホタル生態研究会より、「2007年度調査研究年報」が発行され、その13-28ページに、以下の報告を発表しましたので、お知らせします。
今坂正一(2008)日本産マドボタル属の研究(予報1)−本土のマドボタル属は何種に分けられるのか?−. 2007年度調査研究年報: 13-28.
なお、本文中、♂交尾器が時計回り、あるいは、反時計回りに挿入される部分で、若干の勘違いによる誤りを書いています。
ここでは、印刷された文章を訂正して、掲載したいと思います。
本文のホームページでの公開を許可頂いた陸生ホタル生態研究会の事務局・小俣軍平さんに、お礼を申し上げます。
日本産マドボタル属の研究(予報1)
−本土のマドボタル属は何種に分けられるのか?−
今坂正一
<はじめに>
2006年11月に板当沢ホタル調査団より「日本産ホタル10種の生態研究, 298pp.」が発行されました。私は、2007年夏頃まで、この本についての知識は全くなかったのですが、突然、事務局の小俣軍平さんから対馬のアキマドボタルについての問い合わせがあり、この本の存在を知ることになりました。小俣さんのご厚意で、2-3日後に初めて手に取って内容を知ることが出来たわけです。
到着した本はA4版の大判で、冒頭には、ムネクリイロボタル、カタモンミナミボタル、ゲンジボタル、ヘイケボタル、ヒメボタル、オオマドボタル、クロマドボタル、オオオバボタル、オバボタル、スジグロボタルの10種の卵、幼虫、蛹、雌を含む成虫、生息場所などのカラー写真が掲載されています。ホタル類の幼生期については、形態も含めて、水生のゲンジボタル、ヘイケボタル以外はほとんど知識が無かったので、これほどまで、生態が解ってきたのかと驚きと共に感心した次第です。
事務局によると、「日本のホタルの生態に関する文献は、これまでゲンジボタルとヘイケボタルが中心で、陸生のホタルの生態研究に関するものはすくなく、未解明な点が沢山ありました。また、明らかにされている内容についても、追試がほとんどなされておりませんでした。例えば、? 成虫の発光、? 幼虫の食餌、? 幼虫の棲息場所、? 幼虫の脱皮、? 幼虫の天敵 などがあります。そこで、私たちはずぶの素人集団ですが、室内よりもできるだけ現場に張り付いて追試をしてみようと思いました。そして、その結果をまとめたのが、この本です。ただ、生態研究に関する基礎学習をしっかり積んだ者ばかりではありませんので、見当違いや、思い違いが沢山あることと思います。その点のご指摘を含めて、この本が、日本に生息するホタル、とくに陸生のホタルの生態研究の突破口になれればこんなに嬉しいことはありません。」と述べられています。
板当沢ホタル調査団は、もともと事務局の小俣さんを始めとして、八王子市上恩方町板当沢をフィールドとして、クロマドボタルなど陸生ホタル類の生態を研究しようと始められた会だということです。この調査は、一応、1998年から2001年まで、冬期に生息地の落葉を一枚ずつはぎ取って幼虫を探すような地道で辛抱強い調査を板当沢において続けられ、その間に陸生ホタルの卵・幼虫・蛹の形態を含めて、多くの新たな生態知見を発見されたようです。
その際、クロマドボタル幼虫には斑紋の多型が現れることを発見されました。同時に、他の地域の調査により、幼虫の斑紋パターンには場所により違うタイプも現れることを発見されました。その後、調査を全国規模に拡大されて現在に至っているようです。板当沢にはクロマドボタルだけで、オオマドボタルは生息しないことになっていますが、全国的な幼虫の斑紋の多型調査によって、幼虫ではこの2種の区別ができないこと、オオマドボタルの生息地とされる九州・四国地域でも、クロマドボタルと同様な斑紋多型が見られ、場所ごとに斑紋のパターンが異なることなどが述べられています。
オオマドボタルとクロマドボタルとは、本当に別種か、あるいは亜種か、と言った様々な意見があり、両種の斑紋多型がそれぞれ地域的な差があることから、地史まで踏み込んでの研究に発展しているようです。
また、その点について、ミトコンドリアのDNA遺伝子の解析によるアプローチの一端(鈴木・メイ, 2006)も紹介されています。この本は、そうした、調査団の約8年間の集大成であるようです。
この本を手に取ってから、がぜんホタル類の変異と生態に興味が湧いてきました。そして、より多くの人にこの本のことを知っていただきたいと思い、自身のホームページで紹介しました。
<オバボタル属の♂交尾器>
「日本産ホタル10種の生態研究」では、主として幼生期の生態や個体変異について報告されていて、ホタル類の成虫の分類についてはあまり言及されていないようなので、まず、その点を指摘してみようと思い立ちました。
私自身は、近縁のジョウカイボン科の分類が専門ではありますが、ホタル類の♂交尾器は調べたことはなかったので、実際に解剖して抜き出し検視したのは、実は今回が初めてです。
♂交尾器を比較して明らかになったことの一番目は、「日本産ホタル10種の生態研究」文中、同種説があると書かれていたオバボタル Lucidina biplagiata (Motschulsky)(図2)とオオオバボタル Lucidina accensa Gorham(図3)は、明らかに形態分類上は別種だと確信できたことです。それで、最初にそのことをご報告します。
オバボタル属の図版(図2, 3)をご覧下さい。
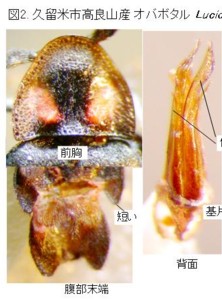
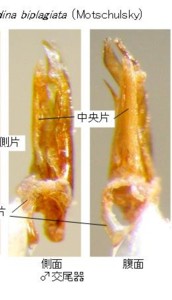
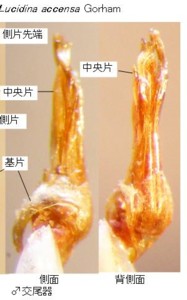
両種共に左上に前胸背、左下に腹部末端節、右側は♂交尾器で、左から背面、側面です。
前胸背、腹部末端節とも、基本構造は一緒ですが、明らかに別種に分けて良い程度の形の違いがあります。
次いで♂交尾器ですが、こちらはさらに違っています。見やすいように、先端部を上にして写真を貼り付けています。
ホタル類の♂交尾器は、甲虫類の♂交尾器の基本形である三叉状の形をしています。図1にすぐ近縁のベニボタル科に属するヒシベニボタル Dictyoptera gorhami (Kono)の♂交尾器を模式的に書いてみました。
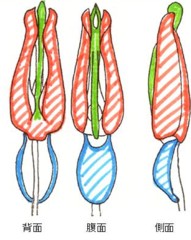

青色で示した部分が基片(basal piece)と呼ばれる部分で、ここから真ん中の緑色で示した中央片(aedeagus、あるいは、狭義のpennis)の中を通って、受精のための射精管が先端まで伸びています。
さらに、この左右両側に1葉ずつ、赤色で示した側片(lateral lobe)が出ていますが、時々、この左右のものが癒着して(この場合parameresと呼びます)筒状になり、中央片を被ったりします。
側片は、普通、交尾の際、中央片を被って保護しながら♀交尾器内に導入し、コメツキなどでは、先端部左右に、外向きのカギ状突起を持っていて、♀交尾器内で広がってイカリを下ろしたように固定し、♀との交接を保つ働きをします。
甲虫の基本形では、全体がほぼ左右対称形で、基片は腹側に位置しています(図1)。
さて、オバボタル属を見ますと、両種とも、基片は、左右非対称で、右側に偏って存在します。
中央片は背面図では側片に隠れて、中央先端部に一部しか見えていません。
側面図では、左側(つまり腹側)に直立しているのが見えると思います。側片は普通左右一対で、オバボタル属では一見基半で左右癒着して半筒状になっているように見えますが、詳細に検討すると、中央でちゃんと左右に分かれています。
先端部分にはそれぞれ側方にトゲ状の突起が見えます。実はオバボタル属の側片は、松葉のように上下に2枚の葉状片が重なった状態になっていて、この突起は、下側の葉状片の先端部が上側の葉状片の脇から顔を出しているものです。
両種共に、この側片の上下2枚の葉状片の先端部はそれぞれ非対称で、互いに違った形をしており、全体として先端は左側に捻れています(細かく云うと捻れの詳細はもっと複雑です)。
(以上の説明の左右は、本来の虫の体の中での話で、後ろ向きの先端を上向きに図示しているので、写真では左右が逆になるものと考えて下さい。以下の説明でも♂交尾器での左右は同様のように見た目と反対になります。)
この捻れについては、後で解説します。
オバボタル(図2)とオオオバボタル(図3)では、側片先端部分のトゲの形や、捻れ具合などが明らかに違うことが解ると思います。
体サイズもまったく違い、さらに、オオオバボタルの♂交尾器は、オバボタルの倍以上の大きさがありますので、交尾は不可能と考えられ、これらは同一種ではなく、形態分類上は明らかに別種です。
<マドボタル属の♂交尾器>
さて、肝心のマドボタル属を後回しにしたのは、こちらの♂交尾器は、オバボタル属のように、形の上で一見して解るような違いが見あたらないからです。
それでも、例として、多少は形態差が解り易い琉球産を先にします。
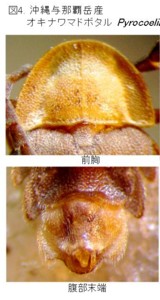
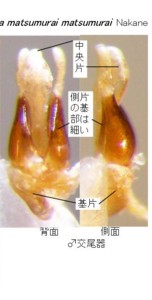
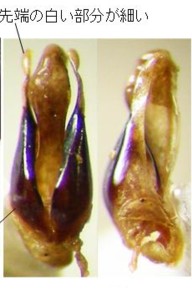
琉球産マドボタル属の図版を見て下さい(図4, 5)。上が沖縄本島与那覇岳産のオキナワマドボタル Pyrocoelia matsumurai matsumurai Nakane (図4)で、
下が西表島カンピラの滝産のサキシママドボタル Pyrocoelia abdominalis Nakane (図5)です。
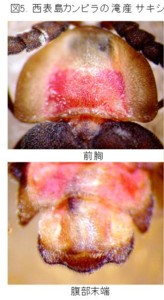
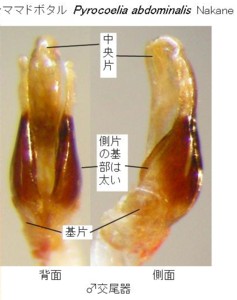
この2種は、大きさ、体色が全然違い、また分布している島も違うので、同じ種と考える人はまずいないと思います。また、色と大きさを度外視しても、前胸背や腹節末端の形でも、比較的簡単に区別できます。
しかし、右側の♂交尾器の写真を見て下さい。オバボタル属とはまったく違っていて、ニンニクの球根みたいな概観をしています。
2種共に形も色もよく似ていて、種の差が顕著ではないことが解ると思います。
左右一対の茶色いラッキョウの球根みたいなところが、先端の白い部分も含めて側片になります。
中央に見える白い部分が中央片です。
オキナワマドボタルの♂交尾器は、側片が中央片とほぼ同じ長さで、褐色部分の基部と先端部の太さの差がそれほど大きくありません。
しかし、サキシママドボタルの♂交尾器は、側片が中央片より短く、褐色部分の基部が特に太く、先端部へ急激に細くなります。
背面図では、前者に対して、後者は全体に縦に引き延ばしたような形をしているので、2種の区別は可能です。
しかし、本質的な形の違いはなく、それぞれの比率が異なるだけなので、写真などに撮って比較しないと、なかなか、見ただけでは判断が難しいかもしれません。
ちょっと見づらいですが、下の方に見える基片に注目して下さい。側面図でも明らかなように、どちらも右半分(写真では左に見える)だけに付いています(基部は半円形に中央片の回りに付着している。)。
左右対称になる通常の甲虫の♂交尾器では、この基片は、中央片の基部腹側に位置しているのが普通です(図1)。それが、オバボタル属同様、マドボタル属でも大部分が右側に移動しているわけです。
ただ、ホタル類の基片・中央片・側片の互いの位置関係は、オバボタル・マドボタル両属共に三叉状で、これは甲虫の♂交尾器の基本的な構造をそのまま残しているわけです。
この琉球の例から考えると、マドボタル属では、みかけがぜんぜん違った種同士でも、♂交尾器の形にはごく微妙な差しか出ないと言うことです。
それで、そのことを念頭に置いた上で、本土のマドボタルの種関係を考える必要があるようです。
逆の言い方をしますと、ごく微妙な差であっても、♂交尾器に一定の差があれば、別種とみなして良いということになるかもしれません。
<ホタル類の交尾姿勢と♂交尾器の形の関係>
オバボタル属では♂交尾器先端が左に捻れ、さらに基片が右側方に移動していました。
マドボタル属では、側片はほぼ左右対称形で、基片のみ右側方に移動していました。これら2つのことは、当然、ホタル類の交尾行動に由来すると考えられますが、どんな理由が考えられるのでしょうか?
この問題の答えは、どんな文献であれ読んだことがなかったのですが、「日本産ホタル10種の生態研究」に掲載されている荻野さんのヘイケボタルの論文(荻野, 2006)を読んで初めて納得しました。
荻野さんは、「雄は雌の背後に近づき、背中にマウントして、その尾端を雌の腹部の側方からあてがって交尾する。やがて雄は雌の背から降り、反対方向に頭を向ける体勢になるのは、ゲンジボタルと同じである。」と書かれています。
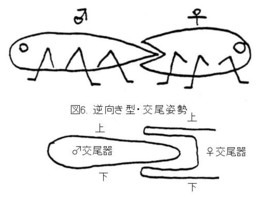
甲虫の交尾姿勢は、より原始的と考えられるグループでは、♂♀がお尻どうしをくっつけ合うようにして交尾し(図6上:逆向き型)、♂交尾器の形は三叉状です。
この体勢では♂交尾器は、背・腹面がそのままの(上下が逆転しない)状態で、♀の交尾器内に挿入されます(マルハナノミ、コメツキ、ジョウカイボン科のMalthodes属など:図6の下)。甲虫以外では、カメムシ類などがこの型として知られています。
一方、甲虫の大部分の進化したグループでは、♂は♀の背面にマウント(図7上:馬乗り型)し、♂交尾器は反転して上下逆さの状態で♀体内に挿入されます(図7下)。
花上で交尾中のカミキリムシとか、ハナムグリ類などはこの形です。
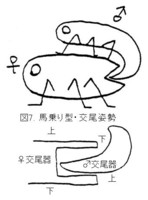
これらの違いは当然、♂交尾器の形の違いに反映され、馬乗り型の甲虫では反転して挿入しやすいように、先端部が腹側に湾曲するものが多く、側片が筒状に変化したり、あるいは側片が退化して中央片がむき出しになり、単純な筒状に変化するものが多いようです。
甲虫以外でも、ハチやハエなどの進化したグループも、ほとんどがこの馬乗り型の交尾姿勢が採用されているようで、交尾の際この姿勢が最も♂が♀の姿勢をコントロールしやすいのではないかと思われます。
しかし、前節で述べたように、ホタル類では逆向き型の♂交尾器の構造を保ちながら、馬乗り型同様の交尾行動をとるので、その逆説的な交尾姿勢に疑問を持っていました。
荻野論文で、その疑問が氷解したわけです。
側方から時計回りに腹端を回して、挿入していたのですね(図8:横から回しバッタ型)。
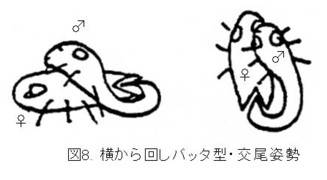
これなら、♂交尾器は上下逆転せずに挿入することができますし、挿入後に♀の背から降りれば、普通の逆向き型の交尾姿勢に戻るわけです。♂交尾器の構造と交尾姿勢の間の矛盾も解消されます。
また、オバボタル属の♂交尾器先端の左への捻れや、オバボタル・マドボタル両属の基片の右側方への移動も納得できます。
交尾器を挿入するためには、先端が左に湾曲している方がうまくいきそうですし、♂交尾器を左から回すためには、基部右側にキチン質の基片が位置する方がより強い力をいれることができると考えられるからです。
この背中にマウントしながら♂交尾器を逆転せずに、側方から挿入する交尾姿勢は、有名な例としては、バッタ類で知られています。
バッタやホタルなど腹部が相当に柔らかく、自由に曲げ伸ばしができるグループでないと、こんな芸当は無理ですね。
横から回しバッタ型の交尾姿勢は、最も原始的と考えられる逆向き型の交尾姿勢からの発展形と考えられます。
実際、ホタル類の♂交尾器はその基本形である三叉状から、ちょっとだけ修正を加えた形でその交尾姿勢に対応しています。
そのことを考えると、根本的に♂交尾器の構造を変化させ、上下を逆転させて対応している馬乗り型の交尾姿勢をとるグループは、相当進化した群と考えることが出来ます。
そう言った意味でも、これらのホタルは、まだ、相当古い形を残していると判断されます。
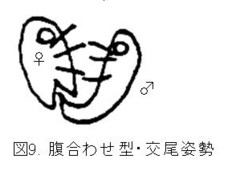
蛇足ながら、チョウ・ガで普通に見られる交尾姿勢(図9. 腹合わせ型)は、逆向き型の変形と考えられ、逆向き型を腹合わせに折り曲げただけの姿勢と考えられます。
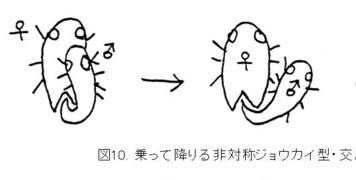
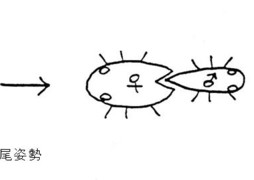
さらに、特殊な交尾姿勢として乗って降りる非対称ジョウカイ型が存在します。
この形は、横から回しバッタ型から、馬乗り型への移行形の交尾姿勢と考えられ、こういう課程を経て、根本的に♂交尾器の構造を変化させて、完全に♂交尾器の上下を逆転させたのであろうと考えています。
その例として、ジョウカイボン科の1亜科であるコバネジョウカイ類は、♂が♀の背面にマウントした状態で挿入を開始し、そのまま右方向へ降りて、最終的に反対向きになります。
そうすると、♂交尾器は上下逆の形からスタートして、♀交尾器内で螺旋状に180度回転すると想像されます(図10. 乗って降りる非対称ジョウカイ型)。
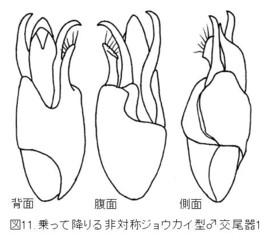
この類の♂交尾器は例外なく左右非対称で、付属の突起を含めて先端部分が時計回りに捻れています{図11. キベリコバネジョウカイ Trypherus niponicus (Lewis)の♂交尾器}。
また、極端な例では、ホタルと近縁であるベニボタル科のミワハナボタルPlateros miwai M. Sato et N. Ohbayashiの♂交尾器(図12. Nakane, 1969より引用)のように、先端部分がワインの栓抜きのように螺旋状に一回転以上しているものもあります。これはもう、♂交尾器を(体も)何回も回転しながらでないと、♀内に挿入することは出来ないだろうと考えています。

この種の場合は今までの全てのグループと異なり、♂交尾器は反時計回りに捻れていますので、マウントした状態で左側に降り、さらに廻って右からマウントし、ということを繰り返すのでしょうか?
グループによって左回り(時計回り)と、右回り(反時計回り)があるというのが興味深いですね。アリヅカムシのある種では、1種の中に、♂交尾器先端が、左回りに捻れている個体と、右回りのとがあるそうです。♀はその両方に対応できるのでしょうか?
<本土のマドボタル類>
♂交尾器の話で、大分脱線しましたね。
解っていただきたかったのは、マドボタル属の♂交尾器が一見普通だけれど、かなり特殊だと言うことと、形に種差が出にくいということでした。
それも、構造をちゃんと把握した上でないと、どのあたりに種の特徴が出るのかを検討することが出来ないと考えたわけです。
オキナワマドボタルとサキシママドボタルの場合は、成虫をパッと見たとしても、色だけでも間違う人はいないと思います。
しかし、♂交尾器の形だけ見せられたら、区別できないかもしれません。
慣れれば、図版に示したように、一定の差は見つかると思いますが・・・。
それでは、本題のオオマドボタル・クロマドボタル群に入ります。
過去に於いては、本土に分布するこの2種は、ほとんど体のサイズと前胸の斑紋で、区別されていたように思います。
そのため、人によっては別種と考えることに疑問を呈し、2種の混棲地や分布範囲などの点でも諸説有って、定説すら良く解らない状態でした。
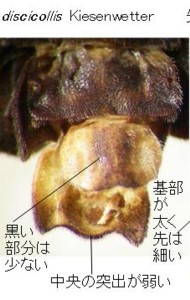
一応、図鑑類では、前胸に赤い斑紋を持つものをオオマドボタル Pyrocoelia discicollis Kiesenwetter、分布は近畿以西、
全体黒色で前胸に斑紋を持たないものをクロマドボタル Pyrocoelia fumosa Gorham、分布は近畿以東ということになっています。
本土のマドボタル属の図版(図13〜16)を見て下さい。
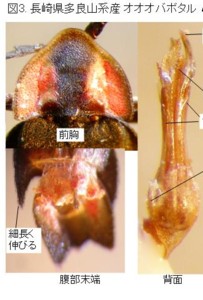
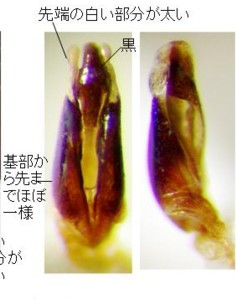
今のところ、私の所の材料は大変少ないのですが、オオマドボタル(原産地は長崎市:Kawashima, Suzuki and M. Sato, 2003より引用)と判断しているのは、やや大型で前胸に赤褐色の紋がある個体で、原産地に近い長崎県雲仙産(図13)と、その周辺の田代原産(図14)です。
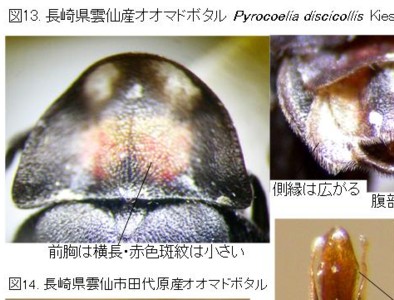

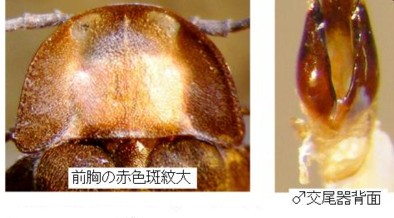

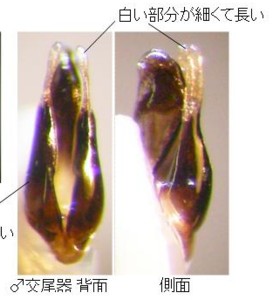
この2産地の♂交尾器は、側片の基部が太く、先へ急激に細まり、先端の白い部分が細長くて、先端の球桿部直前で括れる点など、ほとんど同じ形をしていますので、この2個体は同種(オオマドボタル)と考えられますが、前胸の斑紋の形と大きさにはだいぶ変異があるようです。
さらに、手元にやや小型で、前胸に赤色斑紋がある広島市のもの(図15)があります。


この個体も赤色斑紋が見られるので、従来の考え方ならオオマドボタルに入りますが、若干感じが異なります。
♂交尾器の概観は、雲仙のオオマドボタルとかなり異なり、全体のプロポーションは細長く、黒みが強く(特に中央片の先端半が黒い)、側片基部から先まで比較的同じ太さ、先端の白い部分は雲仙産より、かなり短くなります。
♂交尾器の形態からは、雲仙のオオマドボタルとは明らかに別種程度の差があると判断できます。この種を(仮称)ヒロシママドボタルと呼ぶことにします。
また、滋賀県神津畑(比良山系:図16)のものは全体黒色で、前胸に斑紋が見られないので、一応クロマドボタルの範疇であると考えられます。


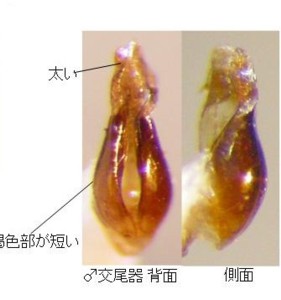
本種の原産地は福島、又は筑波山(Kawashima, Suzuki and M. Sato, 2003より引用)とされているので、現時点では滋賀県産がその地域のものと全く同じかどうか確認できていません。ここではクロマドボタル?としておきます。♂交尾器は前2種とはやはり違っていて、やや薄色の色彩や、側片の基部が太い点などは雲仙のオオマドボタルの方が似ています。しかし、側片先端の白い部分は太くて短く、明らかに前2種とは異なっています。
体長は雲仙産 12.2mm、♂交尾器長 1.7mm (♂交尾器長/体長=0.14)、
田代原産13.5mm、♂交尾器長 1.8mm (♂交尾器長/体長=0.13)、
広島市産 9.8mm、♂交尾器長 1.2mm (♂交尾器長/体長=0.12)、
神津畑産 9.0mm、♂交尾器長 1.1mm (♂交尾器長/体長=0.12)です。
オオマドボタルと判断したものの方が、♂交尾器長/体長の比率は、やや大きいようです。
広島市産は体サイズは小さくてクロマドボタル的、斑紋はオオマドボタル的で、「実際はどっちだ?」と悩ましい気持ちがします。
♂交尾器の比率も含めて小さいので、むしろ、全体としてはクロマドボタル的と言えるかもしれません。しかし、♂交尾器自体の色や形は、この中では最も違って見えます。
当然、♂交尾器の大きさは同一種内では、成虫の体長にかかわらずほぼ一定で、比率的に、小型個体はより大きな♂交尾器を、大型個体はより小さな♂交尾器を持つ傾向があります。
つまり、通常、体長差より♂交尾器の大きさの差の方が、むしろ同一種内では少ないわけです。
同種のどの♀とも交尾できる可能性を増やすためには、可能な限り♂交尾器を平均化した方が、♂にとっては都合が良いわけです。
そのことから考えますと、1.7mmの♂交尾器を持つ雲仙産のオオマドボタルと、1.2mmの広島産のヒロシママドボタルは、多分、交尾が出来ない(つまりは別種)と思われます。
ちょっと前節の琉球産マドボタルに戻りますが、同様に、
オキナワマドボタルは体長6.8mm、♂交尾器長 1.1mm (♂交尾器長/体長=0.16)、サキシママドボタル 9.7mm、♂交尾器長 1.55mm (♂交尾器長/体長=0.16)で、♂交尾器の大きさに差が有りすぎて、多分互いの交尾は無理でしょう。
しかし、沖縄産は本土産より、2種共に比率的には体長に対して♂交尾器が大きいようで、この点では2種は似ているようです。
マドボタル類の♂交尾器に種差がでにくいことから、上記のような少数の検視では、結論めいたことはほとんど言えないようです。
しかし、以上の結果からは、少なくとも長崎県雲仙産、広島市産、滋賀県神津畑産は、それぞれ別の種である可能性が大変大きいことは断言できます。
近畿以西にも、前胸に赤紋があってオオマドボタルではない個体群が生息している、つまりは2種以上分布している可能性が指摘できると思います。
<長崎県多良山系五家原岳山頂の2種のマドボタル>
その後、他の虫を調べる目的で、昔採集した紙包み標本を見直していましたところ、さらに興味深い事実が判明しました。
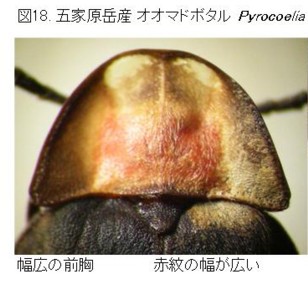
と言いますのは、私のベースグラウンドの1つにしておりました、長崎・佐賀の県境に位置する多良山系で、同日に五家原岳山頂でまったく異なる2種のマドボタルが採れていることに気が付いたのです(図17)。

この事実は、先頃、「ヒメマドボタル(仮称)の発見」と題して、陸生ホタル研の月報 No. 5に掲載していただいた(今坂, 2008b)ので、あるいは、ご存じかと思います。
あまりその記事と重複しない形で要点を述べますと、多良山系の長崎県諫早市五家原岳で、霧と共に風で吹き上げられて来て、山頂付近の葉上に静止中のマドボタルを2種採集しています。
1980年7月15日に、12.8mm程度のもの(オオマドボタル:図18)を1♂、9mmちょうど、くらいの小型のもの(仮称ヒメマドボタル:図19)を1♂、同時に採集していました。
その他に、さらに年度を変えて、同時期に合計オオマドボタル5♂、ヒメマドボタル2♂を採集しています。それぞれの種は、オオマドボタルが12mm以上、ヒメマドボタルが9mm以下で、体長の重複はありません。


♂交尾器の長さは、オオマドボタル1.7mm内外、ヒメマドボタルが1.2mm内外です。
オオマドボタルの方は体長・♂交尾器長共に、長崎県雲仙産の計測値とほとんど同じです。外形等の差も見受けられず、♂交尾器もよく似ているので、この多良五家原岳産は雲仙産と同じく、現在の所、オオマドボタルと判断して良さそうです。
一方、ヒメマドボタルの計測値は、♂交尾器/体長がほぼ0.12で、広島市産のヒロシママドボタル、および、滋賀県神津畑産のクロマドボタル?とほぼ同じです。
一見して気が付くヒメマドボタルの特徴(図19)は、体サイズと♂交尾器が小さいこと以外に、
前胸の形がより縦長であること、
前胸前縁沿いの透けたいわゆる窓の形がくっきりと細く、ハの字が立ってくる(両方の窓の角度が急角度)こと、
四角の赤紋の幅が狭く、中央部の隆起付近で二分されかかっていること、
腹部背板末端節の中央の突出が強く、左右両方の扇型の突起より後方まで突出すること、などです。
いずれの特徴でも、同時に採集されたオオマドボタルとは、明らかに異なりますので、五家原岳山頂では、明らかに2種が共存すると考えて間違いないと思います。
山頂への吹き上げで採集されたことから、山頂を中心とした1〜2kmの範囲に生息していると考えられますので、2種が微環境で棲み分けているのか、あるいはまったく同一環境に生活しているのか、そのどちらかだと思います。
<他産地の小型マドボタルとの比較>
五家原岳産の2種のマドボタルのうち、大型のものは一応、オオマドボタルということで良いとして、では、ヒメマドボタルと仮称した方は、ヒロシママドボタルやクロマドボタル?とどういう関係になるのでしょうか?
先ほどの図13-16を見直してみて下さい。
ちょっと見た感じでは、ヒメマドボタルはヒロシママドボタルに一番よく似ているようです。
特に全体のプロポーション、前胸の色彩、♂交尾器の色合い(中央片の先端半が黒づむこと)や、側片の黒色部が一様に細長く伸びる点は良く似ているようです。
しかしながら、ヒメマドボタルは前胸が縦長になり、腹部背板末端節の中央の突出が強く、側片の白い部分が太くて短くなる点で、ヒロシママドボタルと区別できます。
当然、より似ていないと感じられるオオマドボタルやクロマドボタル?とは、それ以上に種々の差があります。
ということは、現時点での見解として、上記の2個体群と2個体は、全て別種と見なさないと仕方がないと言う結論になります。
さらに、最近鹿児島県で採集されたマドボタルを入手したので、図示してみます(図20)。


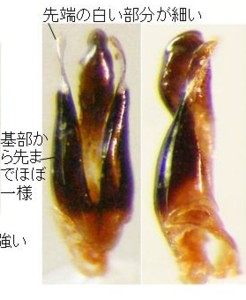
体長は10.5mm、♂交尾器長は1.3mm、♂交尾器/体長はほぼ0.12、今までの小型のマドボタルよりやや大きいですが、比率などから考えても、この小型のタイプに入ると思います。
常識的に考えると、多良山系のヒメマドボタルにより近いのではないかと考えてしまいますが、幅広の前胸、♂交尾器の側片先端の白い部分はむしろ細くて長く、ヒロシママドボタルに最も良く似ています。
しかし、♂交尾器の側片先端の白い部分の形は微妙に異なっていますし、腹部背板末端節の中央の突出が強く、末端節の側縁は直線的になることなど、ヒロシママドボタルと違う点もあります。
またしても、一応、(仮称)カゴシママドボタルと呼ぶことにします。
以上、さらに混沌としてきましたが、各地で少なくとも小型の一群には様々な形をしたものがいると言うことは、解っていただけると思います。
ただし、まだ、検視した個体数が各地点共にごくわずかなので、個体変異の幅が掴めていないと推定され、種の範囲を正しく認識できていない可能性も残されています。
<日本本土産は何種?>
現時点で言えることは、日本本土産のマドボタル属は、
少なくとも大型のものと小型のものの2種以上の種が存在するということ、
そして、小型のものは各地で微妙に変化しているということです。
小型のもののうち、検視できたものは一応、区別可能なので、仮に和名を提唱しましたが、これらが真に種であるか、亜種であるか、それとも、単なる個体変異であるか(この可能性は少ないと思います)、今後、それらを検証する必要があります。
ホタル研の月報等でも、兵庫県と四国において、それぞれ、ほぼ同じ地域に2タイプのマドボタルが見つかった例が報告されていますが、事情は九州とそれほど変わらないのではないかと予想しています。
少なくとも近畿以西では、大型のものと、小型のものの2種が混棲するとみてまず間違いないでしょう。
そのうち、小型のものは各地で変化していますので、将来何種か、あるいは何亜種かに区別されうると推測されます。
一方、大型のものは、まだ、九州以外の個体を検視していないので、地域によって変化しているかどうか予想が付きません。
今後、標本が入手できるようなら、調べてみたいと思っています。
ということで、現時点では、日本本土(本州・四国・九州)産のマドボタル属は、少なくとも1種ではなく、2種以上生息するということしか言えないようです。
<マドボタル属の幼虫研究の問題点>
私のマドボタル属の探求は、「日本産ホタル10種の生態研究」に掲載されている、小俣事務局長を中心とする幼虫の斑紋変異の研究結果を、読ませていただいたところからスタートしました。
そう言う意味で、マドボタル研究上は「原点」ということになります。
その論文の中で、幼虫の斑紋パターンにはさまざまなタイプがあって、地域的に特徴的なものや、出現しないタイプ、普遍的なタイプなどが渾然一体となっており、その複雑なパターンを綿密に整理し、地史ともからめて論じておられます(小俣, 2006)。
しかし、上記のような事実が明らかになってきますと、少なくとも西日本の幼虫の資料の中には、一ヵ所で2種分の斑紋パターンが重複して入っている可能性があります。
同じ採集地点で、2種の幼虫が同時に得られる可能性が高いのか低いのか、今のところ解りませんが、今後、幼虫の斑紋変異を考える際は、その点を計算に入れておく必要があろうかと思います。
先のホタル研の月報4号で、「マドボタル属幼虫の検討(予報)」として、マドボタル幼虫の地域変異について、若干の知見を発表させていただきました(今坂, 2008a)。その中で中間的と書いた日田の幼虫の一部や、伊豆の幼虫の一部には、これら2種の幼虫が混じっていたのかもしれません。
この文章で日田などのオオマドボタルの幼虫の特徴と推定したのは次の通りです。前胸腹面の前縁に、顕著な4つの突起を持つ、
腹節背板の後から2番目の節は中央に三角形(やや小さい)の黒紋を持つ、
前肢脛節は基部1/3以下の狭い範囲が黒色。
これらは、オオマドボタルを含む大型マドボタルの幼虫の特徴ではないかと考えています。
同様に、クロマドボタル幼虫の特徴とした、
前胸腹面の前縁の突起は小さく微弱、
腹節背板の後から2番目の節は中央の黒紋は大きく、側縁が狭く白色、
前肢脛節は基部1/2あるいはそれ以上が黒色、
などの形質は、ヒメマドボタルを含む小型マドボタルの幼虫に共通してみられる特徴の可能性があります。
しかし、地域ごとの種・亜種間の差は、これらの特徴では掴みきれないような気がします。地域個体群ごとの詳細な解明には、あるいは、当初の斑紋パターンが有効かもしれませんので、今後さらに調査を続けて欲しいと期待しています。
一つ気が付いたのですが、幼虫の腹節背板の後から2番目の節の色彩は、側方の白い部分が発光器と考えられ、成虫になってからの末端腹板の発光器の大きさとも対応しているようです。
成虫・幼虫共に、大型マドボタル類で比率的に白色部は大きく、小型マドボタル類で小さくなっているようです。
また、先日、小俣事務局長から五島列島でマドボタル属の幼虫を採集された方を紹介していただきました。その方が五島列島からの幼虫の文献記録を探されていたので、幼虫記録はまだ無いことと、成虫の記録は私自身がとりまとめた文献があることをお知らせしました。
それでようやく、五島に、以前の目で見るとオオマドボタルと判断されるマドボタル属の種が分布することを再認識しました。
残念ながら、記録した個体は共著者の採集品で、手元にないので再検討はできませんでしたが、本土でこれだけ変化していることを考えると、離島ではさらに分化が進んでいることが予想されます。
今後是非、入手して検討してみたいと考えています。
各地の離島、特にさまざま固有種が知られる佐渡、伊豆諸島、隠岐、瀬戸内海の島々、壱岐、五島列島、甑島、屋久島、種子島などでの新発見が期待されるので、今後これらの地域に出かけられる方は注意して探してみて下さい。
引用文献
今坂正一, 2008a. マドボタル属幼虫の検討(予報). 陸生ホタル研 月報, (4): 13-15.
今坂正一, 2008b. ヒメマドボタル(仮称)の発見(予報). 陸生ホタル研 月報, (5): 2-4.
Kawashima, I., Suzuki, H. and M. Sato, 2003. A check-list of Japanese fireflies (Col., Lampyridae and Rhagophthalmidae). Jpn. J. syst. Ent., 9(2): 241-261.
Nakane, T., 1969. Fauna Japonica, Lycidae (Insecta: Coleoptera). 224pp. Biogeographical Society of Japan.
荻野 昭, 2006. (4) ヘイケボタル(Luciola lateralis). 日本産ホタル10種の生態研究: 84-90. 板当沢ホタル調査団.
小俣軍平, 2006. 第1章 本州・四国・九州産マドボタル属幼虫における背板斑紋の変異と地理的分布. 日本産ホタル10種の生態研究: 193-228. 板当沢ホタル調査団.
鈴木浩文・メイ モウ ティ, 2006. オオマドボタルとクロマドボタルをめぐる諸問題とミトコンドリアDNAの解析−途中経過報告−. 日本産ホタル10種の生態研究: 229-234. 板当沢ホタル調査団.

