
このトピックは、2008年6月18日にアップしましたが、2024年のリニューアル時に消失したので、再録しました。
第二番目の種群として、今まで1種とされていた日本のヒメキバネサルハムシが、4種に分割されたことをお知らせします。
この解説を引用したい方は、「佐賀の昆虫」に掲載した、今坂・南(2008)を引用下さい。
今坂正一・南 雅之, 2008. 日本産Pagria(キバネサルハムシ属)について. 佐賀の昆虫, (44): 253-263.
なお、このホームページの解説も、今坂・南の共同制作です。
ヒメキバネサルハムシ Pagria signata (Motschulsky, 1858) は、サルハムシ亜科の中では最も小型のサルハムシの1種(体長1.8-2.8mm程度)で、本州以南~琉球に広く分布しており、各地のマメ科植物にごく普通に見られます。一般に、頭と胸、上翅の合わせ目は黒色、肢と上翅の大部分は黄褐色ですが、個体により、全体ほぼ赤褐色のものから、全体黒色になるものまで、多様な色彩変異があることが知られていました。
つい最近、ロシアの研究者Moseyko氏とMedvedev氏によって、この種とその近縁種についての再検討が行われ(2005a:ロシア語)、次いで英訳版(2005b)も公表されました(文献名は文末の参考文献を参照して下さい)。
Moseykoらはこの論文の中で、P. signataに対してビルマ産の個体を後模式標本として指定し、産地としてベトナム、ラオス、タイ、ミャンマー、インドを上げていますが、日本は分布から除いています。
一方、P. signataのシノニムとなっていた4種を独立種に戻しましたが、その内の3種は過去に日本から新種記載されていた種で、改めてそれらの種の検視データも示し、検索表及び各種の解説で各々の特徴を述べています。
ということは、日本産のヒメキバネサルハムシは3種に別れることになるので、彼等の論文を参考にして、国内産が実際に3種に区別できるのか、それぞれの種がどのような分布をしているのか、調べてみました。
その結果、Moseykoらがウスリーから新種として記載した1種を追加して、国内に4種のPagria (キバネサルハムシ属: 新称)が分布することが明らかになりました。
最初に述べたように、本属の種は普通種と考えられ、各種アセスメント調査や、各地の目録には必ず登場することから、皆さんの注意を喚起する意味で、区別点を明らかにしてここに紹介したいと思います。
<日本産キバネサルハムシ属の区別方法>
日本産4種は、以下の検索表で区別することが出来ます。
なお、従来、ヒメキバネサルハムシという和名で記録されたものは、4種を含んでいますので使用せず、それぞれの種に新たな和名をつけました。
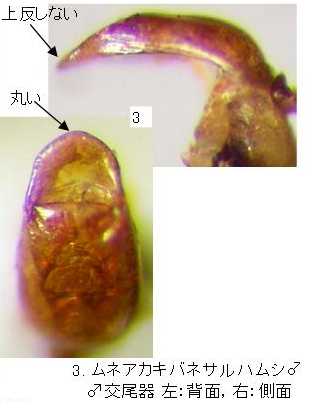
1(2). ♂(写真1)は頭部と前胸背板全体が赤褐色。♀(写真2)はやや黒化し、時に頭頂部のみが赤褐色、前胸背板は黒色で前縁1/3程度は赤褐色。頭頂(写真13)の側縁は複眼の内側でやや角張って波曲する。上翅基部の隆起部の後方に八の字形になる濃色の斑紋を持つが、時に薄まって消失する。♂交尾器(写真3)は先端付近まで太く厚く、前縁は丸く、上反しない。・・・・・・・・・・・・・・・・・・・・ムネアカキバネサルハムシ(新称) Pagria consimile (Baly)
(写真1)

(写真2)

(写真3)
(写真13-14)

2(1). ♂♀共に頭部と前胸背板は大部分が黒色、時に頭頂部と前胸背板前縁は狭く赤褐色。頭頂の側縁は複眼の内側でほぼ直線状(写真14, 15)。♂交尾器は先端部が扁平で、前縁は上反する。
(写真15-16)

3(6). 頭部と前胸背板は黒色。上翅基部側縁は直線状で、ほぼ平行。頭頂の側縁は複眼の内側で直線状。♂交尾器前縁はほぼ切断状で、先端の上反部は広く扁平。
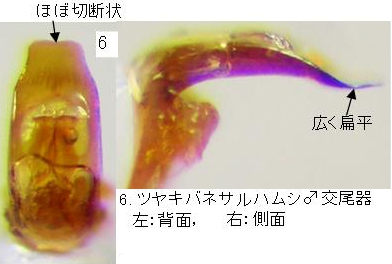
4(5). 上翅はやや細く長い。♂(写真4)の上翅の合わせ目と側縁は黒く、基部の隆起部の後方に黒紋を持たない。♀(写真5)は頭部~上翅まで全体ほぼ全体黒色。前胸背板は光沢が強いが、金属光沢は無い、点刻はより小さく、疎。♂交尾器(写真6)前縁の上反部は広く扁平、中央はややくぼむ。・・・・・・・・・・・・・・・・・・・・・・・・・・・・ツヤキバネサルハムシ(新称) Pagria flavopustulata (Baly)
(写真4)

(写真5)

(写真6)
5(4). 上翅はやや丸く短い(写真7)。合わせ目と側縁は黒く、基部の隆起部の後方は丸く窪み、やや黒づむ。左右の黒紋は繋がらない。前胸背板は金属光沢を持ち、点刻はより強く、密。♂交尾器(写真8)前縁は急角度で上反し、狭い。・・・・・・・・・・・・・・・・・・・・・・・・・・チビキバネサルハムシ(新称) Pagria ingibbosa Pic
(写真7)

(写真8)

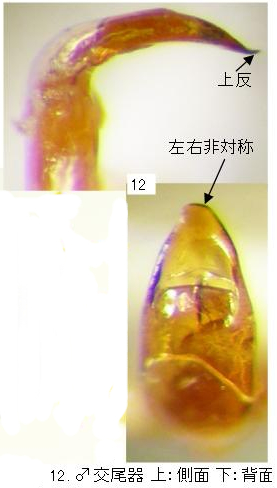
6(3). 頭部と前胸背板は黒色、時に頭頂部と前胸背板前縁は狭く赤褐色。上翅全体に丸みが強く、やや短い(写真9)。頭頂の側縁は複眼の内側で弱く波曲する(写真16)。上翅基部の隆起部の内方~後方に黒紋を持ち、合わせ目を通じて左右の黒紋が菱形あるいは八の字状に繋がる(写真10)。しばしばこの黒紋は褐色に薄まり、時に消失する。頭頂の点刻はやや大きく密。♂交尾器(写真12)先端はやや尖り、微妙に左右非対称。・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・マルキバネサルハムシ(新称) Pagria ussuriensis Moseyko et Medvedev
(写真9)

(写真10)

(写真11)

(写真12)
<各種の解説>
- ムネアカキバネサルハムシ
日本産の本属の中では、最も大きい(2.2-2.8mm)。♂は触角・肢・頭部・前胸背板と、上翅の大部分が赤褐色(写真1)で、上翅の合わせ目は細く濃色。♀は頭頂と前胸背板の前縁中央部1/3程度を除いて黒色(写真2)
上翅基部の隆起部の後方に、八の字形の濃色の斑紋を持つが、時に薄まって消失する。上翅基部側縁も狭く濃色。♂♀共に上翅は比較的長く、前胸背板の点刻は大きく密、上翅の点刻は大きいが浅く、後半は不明瞭。頭頂の側縁は複眼の内側で、やや角張って波曲する(写真13)
♂交尾器(写真3)前縁は丸く、先端は上反しない。
分布:本州、九州、対馬;ロシア、南朝鮮、台湾、インドネシア (ロンボク島、スマトラ)
本州と九州では低地から低山地(標高800m以下)までかなり普遍的に分布。主としてクズやハギ類から採集されています。 - ツヤキバネサルハムシ
体長はやや小さい(2.2-2.4mm)。頭部と前胸背板は黒色で光沢が強い(写真4)。
前胸背板の点刻は小さく疎、♀はやや密。頭頂の点刻は密。頭頂の側縁(写真14)はほぼ直線状、前頭で内側に大きく曲がる。
♂の上翅は黄褐色でやや細長く、基部側縁は平行、合わせ目と側縁のみが黒く、隆起部後方の黒紋を欠く。
♀(写真5)は足を除いてほぼ全体が黒く、上翅基部の隆起部の側方に短い斜めの黄褐色紋を持つ。上翅の点刻は小さいが翅端近くまで明瞭。
♂交尾器(写真6)前縁の上反部は広く扁平、前縁はほぼ切断状で、中央はややくぼむ。
分布:本州、四国、九州、対馬、下甑島;台湾、南中国、ベトナム、タイ、ミャンマー、ネパール、南インド、インドネシア(スマトラ、ジャワ)、フィリピン(ルソン島、ミンドロ島).
国内では一般に局地的で少ないようです。やはりクズから得られています。 - チビキバネサルハムシ
日本産の中で最も小型(1.9-2.3mm)。頭部と前胸背板は黒く、青~銅色の金属光沢を持つ(写真7, 15)。
前胸背板の点刻はやや大きく密、側方はより大きい。
頭頂の点刻(写真15)は小さく疎。頭頂の側縁はほぼ直線状。
上翅は黄褐色で、やや短く丸みが強いが、基部側縁は平行、合わせ目と側縁は黒く、基部の隆起部の後方は丸く窪み、やや黒ずむ、左右の黒紋は繋がらない。上翅の点刻は小さいが中央後まで明瞭。
♂交尾器(写真8)前縁は急角度で上反し、狭い。
分布:本州、九州、壱岐、琉球(阿嘉島);中国南西部、ベトナム、タイ、シンガポール、マレーシア、インドネシア(スンバ島、ロンボク島)、ネパール、ハワイ.
Moseykoらは本州、壱岐から記録し、東南アジアには広く分布するようですが、実際に日本産として標本を確認できたものは、九州南部と阿嘉島などごく少数で、かなり稀な種のようです。 - マルキバネサルハムシ
ムネアカキバネサルハムシに次いで大きい(2.2-2.7mm)。頭部と前胸背板は黒く(写真9)、光沢があるが、時にほとんど光沢のない個体がある。時に頭頂と前胸背板前縁中央が狭く赤褐色。頭部と前胸背板の点刻は大きく密。
頭頂の側縁(写真16)はほぼ直線状でわずかに複眼の内側で波曲する。
上翅は黄褐色(写真10)で、短く、丸みが強い。上翅の合わせ目と側縁は黒く、上翅基部の隆起部の内方~後方に黒紋を持ち、合わせ目を通じて左右の黒紋が菱形、あるいは八の字状に繋がる。しばしばこの黒紋は不明瞭になり、時に消失する。上翅の点刻はやや大きく、中央後では不明瞭。
♀には2型あり、通常は♂と同型(写真10)であるが、時にツヤキバネサルハムシの♀に良く似て、肢を除いてほぼ全体黒く、上翅基部の隆起部あるいは側方に黄褐色紋を持つ黒化型(写真11)が見られる。ツヤキバネサルハムシと同時に得られることもあるので、注意が必要。本種は上翅が短く、前胸背板の点刻が大きく密であり、頭頂の側縁とその付近の形態でも区別できる。
♂交尾器(写真12)前縁はやや尖り、微妙に左右非対称。前縁は狭く上反する。
分布:本州、九州、対馬;ロシア、中国北部、北・南朝鮮.
本種は、河川のクズに多いようです。
九州の九重高原や脊振山など、1000m近い高原まで上がるとクズは見られず、マルバハギに集まっていました。この標高では、ムネアカキバネサルハムシやツヤキバネサルハムシは見つかりませんでした。
今坂の観察では、長崎県島原半島、福岡県久留米市高良山から耳納山地、佐賀市付近、京都市付近、対馬では、それぞれ、チビキバネサルハムシを除く3種がほぼ同時に採集されていますので、多分、この3種は各地で普通に、ほぼ混生していると思われ、注意が必要です。
また、残るチビキバネサルハムシも九州南部では他の種と混生している可能性があり、琉球ではこの種のみが分布しているのではないかと想像しています。
実際に調査採集して、4種の分布の様子をより明確にしていただければと思います。
記録する際には、上記の検索表を参考にしていただき、できれば明確に違いが判る♂交尾器も確認して、記録するようにして下さい。
なお、蛇足ながら、今までヒメキバネサルハムシとされてきた、Pagria signata (Motschulsky, 1858)は、次のような種です。手元にマレーシア産があるので紹介します。
○Pagria signata (Motschulsky, 1858)
概形はムネアカキバネサルに似るが、前胸背板の点刻はより小さく疎(写真17)。
頭頂の側縁(写真19)は複眼上部で角張って強く波曲し、日本産の全ての種と異なる。
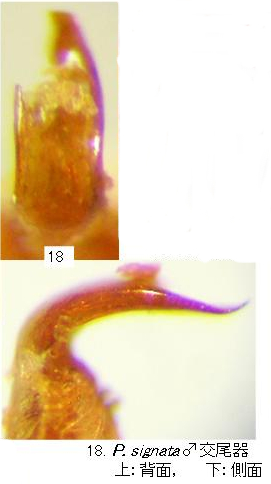
♂交尾器(写真18)はむしろ、ツヤキバネサルハムシに似て、前縁は広く上反し扁平。
図示した個体は、♂交尾器を取り出す際に破損したので、背面からの形は不完全です。
側面からの形は、Moseyko and Medvedev(2005a)の図によく一致します。
(写真17)

(写真18)
(写真19)

参考文献
Baly, J. S., 1867. Phytophaga Malayana. Trans. Ent. Soc. London, 4(2): 77-300.
Baly, J. S., 1874, Catalogue of the phytophagous Coleoptera of Japan, with description of the species new to science. Trans. Ent. Soc. London, 1874: 161-217.
Bryant, G. E., 1942. Notes on the genus Pagria (Eumolpinae, Col.). Bull. Entomol. Res., 33: 31-34.
Chujo, M., 1935. Chrysomelid-beetles of Kyushu belonging to the Umeno Entomological Laboratory. Bull. Umeno Ent. Lab., Kurume, (2): 13-15.
Chujo, M., 1938. Beitrag zur Kenntnis der Chrysomeliden-Fauna von Kyushu, Japan (2). Bull. Umeno Ent. Lab., Kurume, (6): 5-13.
Chujo, M., 1956. A taxonomic study of the Chrysomelidae from Formosa (Insecta: Col.) Part VIII. Subfamily Eumolpinae. Philippine J. Sci., 85(1): 1-180.
Chujo, M. and S. Kimoto, 1961. Systematic catalog of Japanese Chrysomelidae (Col.). Pac. Ins., 3(1): 117-202.
中條道夫, 1959. ハムシ科. 中條道夫ほか著『彦山昆虫目録Ⅱ 鞘翅目』(93pp.): 58-69. 九州大学彦山生物学研究所, 福岡.[Chujo, M., 1959. Family Chrysomelidae. Chujo, M. & al., 『Enumeratio Insectorum Montis Hikosan』(93pp.): 58-69. Hikosan Lab. Biol., Univ. Kyushu.]
Gressitt, J. L. and S. Kimoto, 1961. The Chrysomelidae (Col.) of China and Korea, Part 1. Pac. Ins. Mon. 1A, 299pp.
Kimoto, S., 1961. A revisional note on the type specimens of Japanese Chrysomelidae which are preserved in the museums of Europe and the United States. I. Kontyu, 29: 159-166.
Kimoto, S., 1964. The Chrysomelidae of Japan and the Ryukyu Islands. IV. J. Fac. Agr. Kyushu Univ., 13(2): 235-262.
Kimoto, S., 1969. Notes on the Chrysomelidae from Taiwan II. Esakia, Fukuoka, (7): 1-68.
木元新作, 1980. イネ・ワタ・ダイズのハムシ類害虫-熱帯アジアのハムシ類2. 科学と実験31(3):34-38.
木元新作, 1984. ハムシ科. 林匡夫ほか編著『原色日本甲虫図鑑 (IV)』(438pp.): 147-222. 保育社, 大阪.
Kimoto, S. and J. L. Gressitt, 1966. The Chrysomelidae of the Ryukyu Archpelago. Pac. Ins. 8(2): 467-577.
Kimoto, S. and J. L. Gressitt, 1982. Chrysomelidae (Col.) of Thailand, Cambodia, Laos and Vietnam. III. Eumolpinae. Esakia, Fukuoka, (18): 1-141.
木元新作・滝沢春雄, 1994. 日本産ハムシ類幼虫・成虫分類図説. 539pp. 東海大学出版会, 東京.
木元新作, 2003. タイ・インドシナ産ハムシ類図説. 150pp. 東海大学出版会, 東京.
Jacoby, M., 1908. The Fauna of British India, including ceylon and Burma. Coleoptera. Chrysomelidae. Vol.1, Taylor & Francis, London, 534pp.
Mohamedsaid, M. S., 2004. Catalogue of the Malaysian Chrysomelidae (Insecta: Col.). 239pp. Pensoft, Sofia-Moscow.
Moseyko, A. G. and L. N. Medvedev, 2005a. On the taxonomy of the genus Pagria Lefevre, 1884, with commennts on the genus Rhyparida Baly, 1861 (Col., Chrysomelidae, Eumolpinae). Entomologicheskoe Obozrenie, 84(1): 108-124. [ロシア語版]
Moseyko, A. G. and L. N. Medvedev, 2005b. On the taxonomy of the genus Pagria Lefevre, 1884, with commennts on the genus Rhyparida Baly, 1861 (Col., Chrysomelidae, Eumolpinae). Entomological Review, 85(7): 741-756. [英訳版]
中根猛彦, 1963. ハムシ科. 中根猛彦ほか編著,『原色昆虫大図鑑 II. 甲虫篇』(443pp.): 320-348. 北隆館,東京.
Pic, M., 1929. Nouveautes diverses. Mel. Exot. Ent. 53: 1-36.
Roubal. J., 1929, Coleoptera nova asiatica. Boll. Soc. Entomol. Ital. 61(5-6): 28.
竹中英雄, 1975. ハムシ科. 中根猛彦監修・著, 学研中高生図鑑 昆虫II. 甲虫, 445pp. 学習研究社, 東京.

